Codificación neuronal del sonido
La codificación neuronal del sonido es la representación de la sensación y percepción auditivas en el sistema nervioso. Las complejidades de la neurociencia contemporánea se redefinen de manera continua. Por lo tanto, lo que se conoce del sistema auditivo ha ido cambiando ininterrumpidamente.[1]
La codificación de los sonidos incluye la transducción de las ondas sonoras en impulsos eléctricos (potenciales de acción) a lo largo de las fibras del nervio auditivo, y su posterior procesamiento en el cerebro.
Física básica del sonido
Las ondas sonoras son lo que los físicos llaman ondas longitudinales, que consisten en la propagación de regiones de alta presión (compresión) y regiones correspondientes de baja presión (rarefacción).
Forma de onda
La forma de onda es una descripción de la forma general de la onda sonora. Las formas de onda a veces se describen mediante la suma de sinusoides, a través del análisis de Fourier.
Amplitud
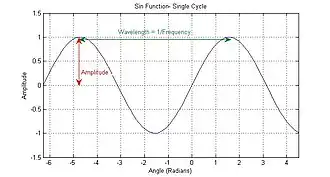
La amplitud es el tamaño (magnitud) de las variaciones de presión en una onda sonora y determina, en especial, el volumen con el que se percibe el sonido. En una función sinusoidal como Csin(2πft), C representa la amplitud de la onda sonora.
Frecuencia y longitud de onda
La frecuencia de un sonido se define como el número de repeticiones de su forma de onda por segundo, y se mide en hercios; la frecuencia es inversamente proporcional a la longitud de onda (en un medio de velocidad de propagación uniforme, como el sonido en el aire). La longitud de onda de un sonido es la distancia entre dos puntos consecutivos coincidentes en la forma de onda. El rango de frecuencia audible para los seres humanos jóvenes está, aproximadamente, entre 20 Hz y 20 kHz. La audición de frecuencias más altas disminuye con la edad y se limita a unos 16 kHz en adultos e incluso a 3 kHz en personas de edad avanzada.
Anatomía del oído
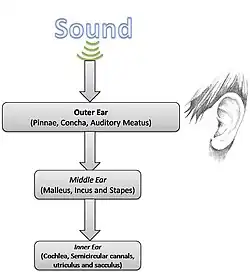
Dada la sencilla física del sonido, la anatomía y la fisiología de la audición pueden estudiarse con mayor detalle.
Oído externo
El oído externo está formado por el pabellón auricular (partes visibles, incluidos los lóbulos y la concha) y el conducto auditivo (el conducto por el que entra el sonido). La función fundamental de esta parte de la oreja es recoger la energía sonora y transmitirla al tímpano. Las resonancias del oído externo aumentan selectivamente la presión acústica con una frecuencia en el rango de 2 a 5 kHz.[2]
El pabellón auricular, como resultado de su estructura asimétrica, es capaz de proporcionar más pistas sobre la elevación desde la que se originó el sonido. La asimetría vertical del pabellón auricular amplifica de forma selectiva los sonidos de mayor frecuencia procedentes de una alta elevación, proporcionando así información espacial en virtud de su diseño mecánico.
Oído medio
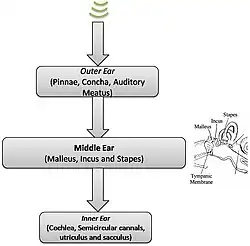
El oído medio cumple una función crucial en el proceso auditivo, ya que, esencialmente, convierte las variaciones de presión en el aire en perturbaciones en los fluidos del oído interno. En otras palabras, es la función de transferencia mecánica la que permite la transferencia eficiente de la energía sonora recogida entre dos medios diferentes. Los tres huesos pequeños que son responsables de este complejo proceso son el martillo, el yunque y el estribo, conocidos en conjunto como los huesecillos del oído. La adaptación de impedancia se desarrolla a través de las proporciones de palanca y la proporción de áreas de la membrana timpánica y la platina del estribo, lo que crea un mecanismo similar al de un transformador. Asimismo, los huesecillos están dispuestos de tal manera que resuenan a 700-800 Hz, al tiempo que protegen el oído interno de una energía excesiva.
Existe un cierto grado de control descendente a nivel del oído medio, máxime a través de dos músculos presentes en esta región anatómica: el tensor del tímpano y el estapedio. Estos dos músculos pueden sostener los huesecillos para reducir la cantidad de energía que se transmite al oído interno en entornos ruidosos.
Oído interno
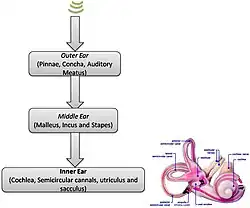
La cóclea del oído interno, una maravilla de la ingeniería fisiológica, actúa como analizador de frecuencias y amplificador acústico no lineal. La cóclea tiene más de 32 000 células ciliadas. Las células ciliadas externas proporcionan, sobre todo, la amplificación de las ondas viajeras inducidas por la energía sonora, mientras que las células ciliadas internas detectan el movimiento de esas ondas y excitan las neuronas (tipo I) del nervio auditivo. El vértice basal de la cóclea, donde entran los sonidos desde el oído medio, codifica el extremo superior del rango de frecuencias audibles, mientras que el vértice apical de la cóclea codifica el extremo inferior del rango de frecuencias.
Esta tonotopía desarrolla una labor clave en la audición, ya que permite la separación espectral de los sonidos. Una sección transversal de la cóclea revela una estructura anatómica con tres conductos principales (rampa vestibular, rampa media y rampa timpánica). En el vértice apical de la cóclea, en una abertura conocida como helicotrema, la rampa vestibular se une a la rampa timpánica. El líquido que se encuentra en estos dos conductos cocleares es la perilinfa, mientras que la rampa media, o conducto coclear, está llena de endolinfa.
Transducción
Células ciliadas auditivas
Las células ciliadas auditivas de la cóclea son el núcleo de la funcionalidad específica del sistema auditivo (en los conductos semicirculares hay células ciliadas similares). Su función principal es la mecanotransducción, o conversión entre señales mecánicas y neuronales.
El número relativamente pequeño de células ciliadas auditivas es sorprendente si se compara con otras células sensoriales, como los bastones y los conos del sistema visual. Por lo tanto, la pérdida de un número menor (alrededor de miles) de células ciliadas auditivas puede ser devastadora, mientras que la pérdida de un número mayor de células de la retina (alrededor de cientos de miles) no será tan grave desde el punto de vista sensorial.
Las células ciliadas cocleares se organizan en células ciliadas internas y externas; interno y externo se refieren a la posición con respecto al eje de la espiral coclear.
Las células ciliadas internas son los principales receptores sensoriales y una cantidad significativa de la información sensorial que llega a la corteza auditiva procede de estas células.
Las células ciliadas externas, por otro lado, potencian la señal mecánica mediante la retroalimentación electromecánica.
Mecanotransducción
La superficie apical de cada célula ciliada coclear contiene un haz de cilios. Cada haz de cilios contiene aproximadamente 300 proyecciones finas conocidas como estereocilios, formadas por elementos del citoesqueleto de la actina.
Los estereocilios de un haz de cilios están dispuestos en múltiples filas de diferentes alturas. Además de los estereocilios, existe una estructura ciliar verdadera conocida como cinocilio, que se cree que juega un rol en la degeneración de las células ciliadas causada por la exposición a altas frecuencias.
Un estereocilio es capaz doblarse en su punto de unión a la superficie apical de la célula ciliada. Los filamentos de actina que forman el núcleo de un estereocilio están altamente interconectados y reticulados con fibrina, y por lo tanto son rígidos e inflexibles en posiciones distintas a la base. Cuando los estereocilios de la fila más alta se desvían en la dirección del estímulo positivo, las filas más cortas también se desvían.
Estas desviaciones simultáneas se producen debido a los filamentos llamados enlaces de punta que unen el lado de cada estereocilio más alto a la parte superior del estereocilio más corto en la fila adyacente. Cuando se desvían los estereocilios más altos, se produce tensión en los enlaces de punta y hace que los estereocilios de las otras filas también se desvíen.
En el extremo inferior de cada enlace de punta hay uno o más canales de transducción mecanoeléctrica (MET, por sus siglas en inglés), que se abren por la tensión en los enlaces de punta. Estos canales MET son canales de transducción selectivos para cationes que permiten que los iones de potasio y calcio entren en la célula ciliada desde la endolinfa que baña su extremo apical. La entrada de cationes, en particular de potasio, a través de los canales MET abiertos hace que el potencial de membrana de la célula ciliada se despolarice. Esta despolarización abre los canales de calcio activados por voltaje para permitir una mayor afluencia de calcio. Esto da lugar a un aumento de la concentración de calcio, lo que desencadena la exocitosis de vesículas de neurotransmisores en las sinapsis en cinta en la superficie basolateral de la célula ciliada. La liberación de neurotransmisores en una sinapsis en cinta, a su vez, genera un potencial de acción en la fibra del nervio auditivo conectada. La hiperpolarización de la célula ciliada, que se produce cuando el potasio sale de la célula, también es importante, pues detiene la afluencia de calcio y, por lo tanto, detiene la fusión de vesículas en las sinapsis en cinta.
Por lo tanto, como en cualquier otra parte del cuerpo, la transducción depende de la concentración y distribución de iones. La perilinfa que se encuentra en la rampa timpánica tiene una baja concentración de potasio, mientras que la endolinfa que se encuentra en la rampa media tiene una alta concentración de potasio y un potencial eléctrico de unos 80 milivoltios en comparación con la perilinfa. La mecanotransducción por estereocilios es altamente sensible y capaz de detectar perturbaciones tan pequeñas como fluctuaciones de fluidos de 0,3 nanómetros, y puede convertir esta estimulación mecánica en un impulso nervioso eléctrico en unos 10 microsegundos.
Fibras nerviosas de la cóclea
Hay dos tipos de neuronas aferentes en el nervio coclear: las de tipo I y las de tipo II. Cada tipo de neurona tiene una selectividad celular específica dentro de la cóclea. El mecanismo que determina la selectividad de cada tipo de neurona para una célula ciliada específica ha sido propuesto por dos teorías antagónicas en neurociencia, conocidas como la hipótesis de la instrucción periférica y la hipótesis de la instrucción autónoma celular. La hipótesis de la instrucción periférica afirma que la diferenciación fenotípica entre las dos neuronas no se produce hasta que estas neuronas indiferenciadas se unen a las células ciliadas, que, a su vez, dictarán la vía de diferenciación.
La hipótesis de la autodiferenciación celular afirma que la distinción en neuronas de tipo I y tipo II se produce después de la última fase de la división mitótica, pero antes de las inervaciones. Ambos tipos de neuronas participan en la codificación del sonido para su transmisión al cerebro.
Neuronas de tipo I
Las neuronas de tipo I inervan las células ciliadas internas. Existe una convergencia significativamente mayor de este tipo de neurona hacia el extremo basal en comparación con el extremo apical. Un haz de fibras radiales actúa como intermediario entre las neuronas de tipo I y las células ciliadas internas. La proporción de inervación que se observa entre las neuronas de tipo I y las células ciliadas internas es de 1:1, lo que da como resultado una alta fidelidad y resolución de la transmisión de la señal.
Neuronas de tipo II
Por otro lado, las neuronas de tipo II inervan las células ciliadas externas. Sin embargo, existe una convergencia notablemente mayor de este tipo de neurona hacia el extremo apical en comparación con el extremo basal. Se observa una relación de inervación de 1:30-60 entre las neuronas de tipo II y las células ciliadas externas, lo que a su vez hace que estas neuronas sean ideales para la retroalimentación electromecánica.
Las neuronas de tipo II pueden manipularse a nivel fisiológico para inervar las células ciliadas internas, siempre que las células ciliadas externas hayan sido destruidas por daños mecánicos o químicos inducidos por fármacos como la gentamicina.
Tronco encefálico y mesencéfalo
El sistema nervioso auditivo incluye muchas etapas de procesamiento de información entre el oído y la corteza.
Corteza auditiva
Las neuronas auditivas primarias transportan potenciales de acción desde la cóclea hacia la vía de transmisión que se muestra en la imagen adyacente. Múltiples estaciones de retransmisión actúan como centros de integración y procesamiento.
Las señales llegan al primer nivel de procesamiento cortical en la corteza auditiva primaria (A1), en la giro temporal superior del lóbulo temporal. La mayoría de las áreas, hasta la A1 inclusive, están mapeadas de forma tonotópica (es decir, las frecuencias se mantienen en un orden establecido). En cambio, la A1 participa en la codificación de aspectos más complejos y abstractos de los estímulos auditivos sin codificar bien el contenido de frecuencias, incluida la presencia de un sonido distinto o sus ecos. Al igual que las regiones inferiores, esta región del cerebro tiene neuronas sensibles a combinaciones que tienen respuestas no lineales a los estímulos.
Estudios recientes realizados en murciélagos y otros mamíferos han revelado que la capacidad de procesar e interpretar la modulación en frecuencias se produce principalmente en los giros temporal superior y medio del lóbulo temporal.
La lateralización de la función cerebral se da en la corteza, con el procesamiento del habla en el hemisferio cerebral izquierdo y los sonidos ambientales en el hemisferio derecho de la corteza auditiva. La música, con su influencia en las emociones, también se procesa en el hemisferio derecho de la corteza auditiva. Aunque no se entiende muy bien la razón de dicha localización, la lateralización, en este caso, no implica exclusividad, ya que ambos hemisferios participan en el procesamiento, pero uno suele desempeñar un papel más importante que el otro.
Ideas recientes
Se ha observado una alternancia en los mecanismos de codificación a medida que se avanza por la corteza auditiva. La codificación pasa de respuestas sincrónicas en el núcleo coclear a depender, a posteriori, de la codificación basada en la tasa de disparo en el colículo inferior. A pesar de los avances en terapia génica que permiten alterar la expresión de genes que afectan a la audición, como el ATOH1, y del uso de vectores virales con ese fin, las complejidades micromecánicas y neuronales que rodean a las células ciliadas del oído interno hacen que la regeneración artificial in vitro siga siendo una realidad lejana.
Según estudios recientes, la corteza auditiva podría no estar tan implicada en el procesamiento descendente como se suponía. En estudios llevados a cabo en primates para tareas que requerían la discriminación de la fluctuación acústica, Lemus descubrió que la corteza auditiva tenía solo una incidencia sensorial y no tenía nada que ver con la cognición de la tarea en cuestión. Debido a la presencia de los mapas tonotópicos en la corteza auditiva en edad temprana, se ha asumido que la reorganización cortical tuvo poco que ver con el establecimiento de estos mapas, pero están sujetos a la plasticidad. La corteza parece efectuar un procesamiento más complejo que el análisis espectral o incluso el análisis espectro-temporal.
Véase también
Referencias
- ↑ Leonard, Matthew K.; Gwilliams, Laura; Sellers, Kristin K.; Chung, Jason E.; Xu, Duo; Mischler, Gavin; Mesgarani, Nima; Welkenhuysen, Marleen; Dutta, Barundeb; Chang, Edward F. (2024-). «"Large-scale single-neuron speech sound encoding across the depth of human cortex"». Nature. 626 (7999): 593–602. ISSN 0028-0836. PMC 10866713. doi:10.1038/s41586-023-06839-2.
- ↑ Hudspeth, AJ. (1989). «"How the ear's works work".». Nature. 341 (6241): 397-404. PMID 2677742. doi:10.1038/341397a0.
Hudspeth, A. J. (2001). «How the ear's works work: mechanoelectrical transduction and amplification by hair cells of the internal ear». Harvey Lectures 97: 41-54. PMID 14562516.
Hudde, H.; Weistenhofer, C. (2006). «Key features of the human middle ear». ORL J Otorhinolaryngol Relat Spec 68 (6): 324-328. PMID 17065824. S2CID 42550955. doi:10.1159/000095274.
Hudspeth, A. J.; Konishi, M. (octubre de 2000). «Auditory neuroscience: development, transduction, and integration». Proceedings of the National Academy of Sciences of the United States of America 97 (22): 11690-11691. PMC 34336. PMID 11050196. doi:10.1073/pnas.97.22.11690.
Kaas, J. H.; Hackett, T. A.; Tramo, M. J. (abril de 1999). «Auditory processing in primate cerebral cortex». Current Opinion in Neurobiology (PDF) 9 (2): 164-170. PMID 10322185. S2CID 22984374. doi:10.1016/S0959-4388(99)80022-1.
Fettiplace, R.; Hackney, C. M. (enero de 2006). «The sensory and motor roles of auditory hair cells». Nature Reviews Neuroscience 7 (1): 19-29. PMID 16371947. S2CID 10155096. doi:10.1038/nrn1828.
Beurg, M.; Fettiplace, R.; Nam, J. H.; Ricci, A. J. (mayo de 2009). «Localization of inner hair cell mechanotransducer channels using high-speed calcium imaging». Nature Neuroscience 12 (5): 553-558. PMC 2712647. PMID 19330002. doi:10.1038/nn.2295.
Rubel, E. W.; Fritzsch, B. (2002). «Auditory system development: primary auditory neurons and their targets». Annual Review of Neuroscience 25: 51-101. PMID 12052904. doi:10.1146/annurev.neuro.25.112701.142849.
Chechik, Gal; Nelken, I. (2012). «Auditory abstraction from spectro-temporal features to coding auditory entities». Proceedings of the National Academy of Sciences of the United States of America 109 (44): 18968-18973. Bibcode:2012PNAS..10918968C. PMC 3503225. PMID 23112145. doi:10.1073/pnas.1111242109.
Frisina, R. D. (agosto de 2001). «Subcortical neural coding mechanisms for auditory temporal processing». Hearing Research 158 (1–2): 1-27. PMID 11506933. S2CID 36727875. doi:10.1016/S0378-5955(01)00296-9.
Brigande, J. V.; Heller, S. (junio de 2009). «Quo vadis, hair cell regeneration?». Nature Neuroscience 12 (6): 679-685. PMC 2875075. PMID 19471265. doi:10.1038/nn.2311.
Lemus, L.; Hernández, A.; Romo, R. (junio de 2009). «Neural codes for perceptual discrimination of acoustic flutter in the primate auditory cortex». Proceedings of the National Academy of Sciences of the United States of America 106 (23): 9471-9476. Bibcode:2009PNAS..106.9471L. PMC 2684844. PMID 19458263. doi:10.1073/pnas.0904066106.
Kandler, K.; Clause, A.; Noh, J. (junio de 2009). «Tonotopic reorganization of developing auditory brainstem circuits». Nature Neuroscience 12 (6): 711-717. PMC 2780022. PMID 19471270. doi:10.1038/nn.2332.
Enlaces externos
- Esta obra contiene una traducción completa derivada de «Neural encoding of sound » de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.