Proceso de hominización
Izquierda: Australopithecus afarensis (3,6 Ma)
Centro: Homo erectus (1,5 Ma)
Derecha: Homo sapiens (actual).
Hominización o antropogénesis es el conjunto de procesos que transformaron progresivamente un linaje de primates en humanos. El proceso evolutivo biológico concernió al grupo de los homininos (Hominina) a partir de la divergencia entre el último ancestro común de los grandes simios (monos antropomorfos, que forman junto como los homininos el grupo de los homínidos —Hominidae—), hace más de cinco millones de años,[1] lo que se conoce como evolución humana.
El término y el concepto que designa fueron acuñados por Édouard Le Roy en su obra Les origines humaines et l'évolution de l'intelligence (Traducido como:Orígenes humanos y la evolución de la inteligencia), publicación de un curso impartido en el Collège de France entre 1928 y 1929. Con anterioridad el término había sido utilizado en un texto inédito de Pierre Teilhard de Chardin en 1924.
Como proceso de evolución cultural no se restringe a cambios anatómicos o fisiológicos, sino a cambios etológicos o conductuales, no innatos sino culturales, que se produjeron y transmitieron paralelamente, tanto en la cultura material (cuya parte más conocidas son las herramientas provenientes de la talla lítica —industria lítica—) como en el lenguaje, la organización social, las mentalidades, las tradiciones, la producción intelectual (técnicas e ideas precientíficas y pretecnológicas, creencias, creación artística) y todo tipo de formas de relacionarse entre sí y con el medio natural (cuyo estudio es más especulativo, al basarse en inferencias y deducciones a partir de los restos materiales y las conclusiones que pueden extraerse de los estudios antropológicos).

Cambios anatómicos
Según diversos estudios paleoantropológicos, el proceso evolutivo de los homínidos incluyó modificaciones anatómicas significativas, como el desarrollo de la bipedestación, [4] un incremento en la capacidad craneal (encefalización)[5] y adaptaciones a cambios ambientales.[6] Estas transformaciones se consideran clave para diferenciar a los homínidos de otros primates.[7][8]
Grandes cambios en el esqueleto
Los cambios en el esqueleto se explican fundamentalmente por el cambio de la postura corporal dominante a la denominada postura o posición erguida o erecta (ortostasis u ortostatismo —vulgarmente, estar "de pie"—)[9] y el bipedismo; cuya causa suele relacionarse con el cambio de hábitat de los primeros hominidos (de un entorno selvático a un entorno de sabana).
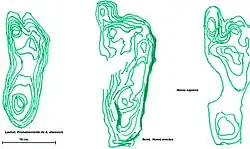
- Las extremidades inferiores se alargan y el pie se hace plantígrado (se apoya toda la planta). El dedo gordo se sitúa en el mismo plano que los otros cuatro dedos.
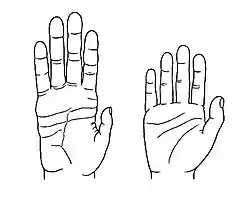
- Las extremidades superiores se alargan y quedan libres, lo que favoreció la manipulación de objetos y el desarrollo de la pinza del pulgar. La versatilidad de la mano humana es una condición fundamental.
- El cráneo se inserta por su base en la columna vertebral, que a su vez adquiere una forma en "S".
- La pelvis se acorta y se robustece, lo cual permite una mayor sujeción de los músculos glúteos, necesarios para caminar erguido.
Evolución del tamaño corporal en humanos
El aumento del tamaño corporal en los homínidos a lo largo de la evolución está relacionado con múltiples factores ecológicos y adaptativos, siendo la disponibilidad de recursos alimenticios uno de ellos. Si bien es cierto que el desarrollo tecnológico (como herramientas de caza y procesamiento de alimentos) permitió un mayor acceso a nutrientes,[10] otros elementos clave incluyen:
- Cambios metabólicos: El aumento del cerebro requirió mayor energía, favoreciendo cuerpos más grandes.[11]
- Presiones selectivas: La termorregulación en diferentes climas influyó en la masa corporal, dando lugar a cuerpos más robustos en latitudes altas (Regla de Bergmann).[12]
- Competencia interespecífica: La necesidad de defensa contra depredadores.
Mientras que Australopithecus promedio medía ~1.2-1.4m, Homo erectus alcanzaba ~1.6-1.8m, coincidiendo con:[13]
- Dominio del fuego (~1.5 Ma).
- Mejoras en técnicas de caza.
- Expansión a nuevos ecosistemas.
| Especie | Estatura promedio | Periodo | Innovaciones asociadas |
|---|---|---|---|
| H. sapiens | 1.5-1.8 m | >0.3 Ma | Tecnología lítica avanzada |
| H. erectus | 1.6-1.8 m | 1.9-0.1 Ma | Uso sistemático del fuego |
| A. afarensis | 1.2-1.4 m | 3.9-2.9 Ma | Bipedestación completa |
Evolución del dimorfismo sexual en homininos
El dimorfismo sexual en estatura ha disminuido significativamente a lo largo de la evolución de los homininos. Los primeros homininos, como Australopithecus afarensis, mostraban un marcado dimorfismo (machos ~30-50% más grandes), similar al de los grandes simios actuales, lo que sugiere una intensa competencia entre machos.[13] Con la aparición de Homo erectus, esta diferencia se redujo notablemente (~15-20%), reflejando cambios en la estructura social hacia sistemas más cooperativos.[14][15]
En humanos modernos (Homo sapiens), el dimorfismo es moderado (hombres ~7-15% más altos), resultado de presiones evolutivas hacia una menor competencia física entre machos y adaptaciones a estrategias de subsistencia cooperativas.[16] Aunque en épocas recientes la brecha ha disminuido ligeramente en algunas poblaciones debido a mejoras nutricionales,[17] la reducción más drástica ocurrió durante la transición de Australopithecus a Homo, vinculada a cambios en el comportamiento social y reproductivo.[14]
-
![Ilustración del libro de Thomas Henry Huxley Evidence as to man's place in Nature.[18]](./Frontispiece_to_Huxley_'Evidence_as_to_man's_place...'_Wellcome_L0063032.jpg) Ilustración del libro de Thomas Henry Huxley Evidence as to man's place in Nature.[18]
Ilustración del libro de Thomas Henry Huxley Evidence as to man's place in Nature.[18] -
 Ídem.
Ídem. -
 Ídem.
Ídem.
Aumento de la capacidad craneal
La expansión progresiva de la capacidad craneoencefálica constituye uno de los hitos evolutivos más significativos en la radiación de los homínidos, correlacionándose directamente con el desarrollo de capacidades neurocognitivas sin precedentes en el reino animal. Este fenómeno biocultural incluye:[19][20]
- Lenguaje simbólico complejo.
- Planificación estratégica a largo plazo.
- Tecnología lítica acumulativa.[7]
Documentado paleoneurológicamente desde el Plioceno tardío (~3.9 Ma) hasta el Holoceno, este proceso manifiesta una presión selectiva multifactorial hacia la encefalización, particularmente marcada en el género Homo, donde el volumen endocraneal experimentó:[21]
- Un incremento del 300% respecto a Australopithecus.
- Una reorganización neuroanatómica cualitativa.
La selección natural favoreció este costoso tejido nervioso (consumiendo el 20% de la energía basal) debido a su valor adaptativo en entornos sociales y ecológicos complejos.
| Taxón | Biocronología (Ma) | Comentarios | Capacidad craneal (cm³) | Cráneo |
|---|---|---|---|---|
| Homo sapiens | ~0,3-0,0 | El hueso frontal se desarrolla haciéndose más vertical, con lo que aparece la característica frente humana, el torus supraorbital se reduce y los lóbulos parietales y frontales se expanden.[22] | 1300–1500 |  |
| H. ergaster H. erectus |
1,9-0,1 | Desarrollo de un hueso frontal más vertical, aunque aún con torus supraorbital prominente.[22] | 800–1100 |  |
| Homo habilis | 2,4-1,4 | Primeros miembros del género Homo. Primeras evidencias de un ligero aumento en el lóbulo frontal, relacionado con herramientas líticas.[23] | 500–700 | 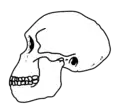 |
| Australopithecus | 3,9-2,0 | Morfología similar a los grandes simios, con un hueso frontal inclinado y ausencia de frente vertical. | 340–500 |  |
-

-
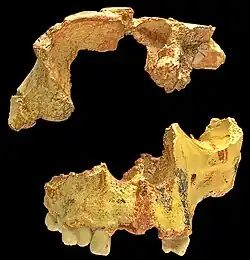 Fragmento de cráneo de Homo antecessor.
Fragmento de cráneo de Homo antecessor. -
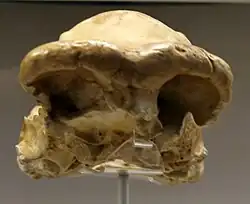 Fragmento de cráneo de Homo ergaster u Homo erectus.
Fragmento de cráneo de Homo ergaster u Homo erectus. -

-
 Comparación de los cráneos de Homo neanderthalensis y Homo sapiens.
Comparación de los cráneos de Homo neanderthalensis y Homo sapiens.
Implicaciones evolutivas
- Encefalización: Aumento en la relación entre el tamaño del cerebro y la masa corporal, medida mediante el cociente de encefalización (EQ). Los humanos modernos tienen un EQ de alrededor de 5 (frente a 2,5 en chimpancés).
- Reorganización cerebral: Expansión de áreas asociadas al lenguaje (lóbulo temporal) y funciones ejecutivas (corteza prefrontal).
- Restricciones obstétricas: El parto humano se ve limitado por el canal pélvico estrecho (producto de la bipedación), lo que favorece un crecimiento cerebral postnatal acelerado.[22]
Este desarrollo no fue lineal, sino el resultado de presiones adaptativas multifactoriales (dieta, sociabilidad, tecnología) para optimizar el balance entre:
- La adquisición de capacidades cognitivas avanzadas (inteligencia).
- Las demandas metabólicas de un tejido neural energéticamente costoso.
- Las restricciones anatómicas impuestas por la biomecánica bípeda.
Este sofisticado balance adaptativo culminó en la singular complejidad neurosimbólica que caracteriza a Homo sapiens, manifestada en pensamiento abstracto, lenguaje articulado, conducta simbólica (arte, rituales) y planificación a largo plazo.
Neotenia
Frente a otras especies, los humanos desarrollan una marcada retención de características juveniles en la etapa adulta (neotenia) y una infancia prolongada, que probablemente está relacionada con la gran plasticidad cerebral.[24]
Los humanos presentan rasgos neoténicos, como:
- Cráneo esferoidal, similar al de una cría de chimpancé, pero que en Homo sapiens persiste en adultos.
- Cara corta y plana, con menos prognatismo que otros primates.
- Dependencia prolongada. Los humanos nacen altriciales y tardan años en ser autosuficientes, a diferencia de otros mamíferos.
Estos rasgos sugieren que, evolutivamente, se ha priorizado el desarrollo cerebral sobre la maduración física rápida. En términos estadísticos el tamaño del cerebro humano en los neonatos representa solo una cuarta parte (25.77%) del tamaño del cerebro de un adulto, mientras que en los chimpancés es un 35.55% y en los orangutanes un 37.61%.[25]
Pérdida de vello corporal
El vello corporal disminuyó drásticamente durante la evolución humana, reduciéndose principalmente al cabello de la cabeza (que crece de forma continua) y al vello en zonas que se desarrollan como caracteres sexuales secundarios a partir de la pubertad (vello púbico, axilas y en algunos grupos, barba). Esta reducción se interpreta como una adaptación clave en entornos de sabana abierta, donde la termorregulación eficiente fue crítica. La pérdida de pelo denso, combinada con un aumento de glándulas sudoríparas, optimizó la disipación de calor durante actividades de esfuerzos prolongados como la caza por persistencia.[26][27] Simultáneamente, la menor densidad pilosa redujo la infestación por ectoparásitos (como piojos y ácaros), minimizando riesgos de enfermedades infecciosas.[28][29]
Estudios genómicos sugieren que la pérdida del pelo se consolidó hace ~1.2 Ma en África, coincidiendo con el uso habitual de refugios que incrementaron la exposición a parásitos.[30] Posteriormente, tras la expansión de Homo erectus a latitudes septentrionales, la especie se adaptó cambiando el color de la piel,[31] no recuperando el vello.[30] Otras hipótesis incluyen la selección sexual,[32] donde la piel visible pudo servir como indicador de salud.[33] Aunque morfológicamente el ser humano parece "desnudo",[34] conserva folículos pilosos en densidad similar a los chimpancés.[35][36] La diferencia radica en que la mayor parte del pelo corporal es fino y corto, excepto en las áreas mencionadas.[37][38][30]
Evolución de la dieta: implicaciones cognitivas y morfológicas
Los cambios paleoclimáticos ocurridos durante el Zancliense y el Piacenziense (3.6-1.5 Ma), caracterizados por una progresiva aridificación del continente africano y la expansión de ecosistemas de sabana, impulsaron una significativa transición dietética en los homininos.[39] Mientras que sus antepasados directos, los Australopithecus, mantenían una dieta predominantemente herbívora y frugívora,[40] basada en frutos, hojas y tallos, los primeros representantes del género Homo (H. abilis y H. erectus) desarrollaron un patrón alimenticio marcadamente omnívoro y oportunista.[41]
Esta adaptación a una dieta más variable e impredecible, que incluía un mayor consumo de recursos animales (carroña, caza menor) y vegetales subterráneos, requirió el desarrollo de capacidades cognitivas avanzadas:[42]
- Planificación compleja: Necesaria para la fabricación y uso sistemático de herramientas líticas (Industria Olduvayense), organización de partidas de caza y recolección y procesamiento de alimentos.
- Memoria espacial ampliada: Fundamental para recordar la ubicación de recursos dispersos, rutas migratorias de animales y fuentes de materias primas para herramientas.
- Cooperación social: Imprescindible para actividades grupales como la caza mayor y la defensa contra depredadores.
- Uso y dominio del fuego, que redujo el esfuerzo masticatorio.
Estos desafíos favorecieron la selección natural de cerebros más grandes y complejos. El aumento del cociente de encefalización requirió, a su vez, una infancia prolongada, que permitió una extensión del aprendizaje y la asimilación de habilidades técnicas y sociales.[43] Esta combinación de flexibilidad dietética, innovación tecnológica y desarrollo cerebral prolongado sentó las bases del éxito adaptativo del linaje humano.[44]

.JPG)
.jpg)
Desarrollo del lenguaje
El lenguaje humano permitió una mejora trascendental en la coordinación de los grupos y la transmisión de conocimientos, disparando la capacidad de evolución cultural y posibilitando el inicio del pensamiento simbólico.[45]
- El desarrollo progresivo del área de Broca, una parte del cerebro relacionada con el habla. Los Homo ergaster y Homo heidelbergensis usaban el lenguaje pero de forma diferente.
- La modificación en la posición de la laringe, la cual en el género Homo ocupó una posición más baja que en el resto de los primates.
- En la Sima de los Huesos del yacimiento de Atapuerca se han encontrado restos tan completos de Homo heidelbergensis que se puede reconstruir hasta el oído medio, de íntima conexión con la capacidad de comunicarse con sonidos.[46]
El fuego
El control del fuego fue una innovación tecnológica crítica en la evolución humana, permitiendo a los homininos paleolíticos obtener calor, protegerse de depredadores y cocinar alimentos. Su dominio transformó patrones sociales y biológicos.[47] Las evidencias sugieren que su manejo fue un proceso escalonado:[48][49][50][51]
- Fase oportunista (~1.5-1 Ma): Uso de incendios naturales (rayos, volcanes) para obtener brasas, sin capacidad de reignición.
- Fase de mantenimiento (~1-0.8 Ma): Conservación prolongada de brasas en "guardafuegos" (recipientes de piedra o hueso), como sugieren restos arqueológicos en Zhoukoudian y la Cueva Wonderwerk en Sudáfrica, donde se encuentra la evidencia más antigua de fuego usado para procesar alimentos (~1 Ma):[52] homininos, probablemente Homo erectus,[53] procesaron restos faunísticos, según análisis de marcas de corte en huesos con signos de calcinación y microestratigrafía de cenizas. Se desconoce si el fuego fue obtenido deliberadamente o si se aprovechó de fuentes naturales.[54]
- Fase de ignición deliberada (>0.8 Ma): Desarrollo de técnicas para crear fuego ex novo (fricción, percusión mineral), documentado en sitios neandertales como Abric Romaní (España). La socialización del fuego está atestiguada desde hace ~0.8 Ma en el yacimiento de Gesher Benot Ya'aqov en el Vado de Jacob.[55][56]
A partir del Paleolítico Superior (~0.03 Ma) en Eurasia se intensificó la expansión de poblaciones humanas hacia latitudes septentrionales y entornos más fríos, donde la movilidad estacional se volvió crucial para la subsistencia.[57] La capacidad de transportar fuego se convirtió en un factor clave para la supervivencia durante estos desplazamientos. Para garantizar su disponibilidad, se emplearon tres métodos principales para producir fuego:[49]
- Taladro de arco (con variantes como arados y sierras): fricción sostenida sobre yesca.
- Pistones de fuego: aprovecha la compresión adiabática del aire en un cilindro de madera y un pistón.
- Encendedor de chispa: percusión de materiales piritosos contra sílex.[58]
Estas tecnologías muestran paralelismos con herramientas paleolíticas de Homo sapiens, sugiriendo un origen común y distribución vinculada a los patrones de migración humana.
Impactos biológicos y sociales
Una vez controlado, el fuego supuso un hito tecnológico que redefinió la ecología humana y generó transformaciones clave:[53][47][50][59]
- Biológicas: la cocción aumentó la absorción de nutrientes, como almidones y proteínas, facilitando el desarrollo del cerebro y reduciendo el tamaño del aparato digestivo.[60][61]
- Expansión geográfica:[62] colonización de regiones frías de Eurasia.[49]
- Sociales: las fogatas actuaron como espacios de cohesión social para compartir alimentos, transmitir conocimientos y desarrollar comunicación simbólica. Se extendió la actividad a horas nocturnas mediante iluminación artificial.
La monogamia
La monogamia es el sistema de emparejamiento predominante en el ser humano moderno (Homo sapiens), incluso en sociedades que permiten ciertas formas de poligamia.[63] A diferencia de otros primates, este rasgo ha sido objeto de debate en la paleoantropología, con hipótesis que sugieren su aparición temprana en el como adaptación con ventajas evolutivas, como la inversión parental y la reducción de conflictos sociales.[64]
Algunos estudios vinculan su desarrollo con la bipedestación y cambios en la estructura social de los homininos, aunque su origen exacto sigue discutido.[65]
Referencias
- ↑ Jaiswal, Ajeet. The hominization process of Homo Sapiens, University of Delhi, India, 2007, p. 43.
- ↑ Guijarro-Clarke, Cristina. «Evolution: that famous 'march of progress' image is just wrong». The Conversation (en inglés). Consultado el 11 de febrero de 2023.
- ↑ Cornelius, Keridwen (16 de agosto de 2022). «The Problems of Evolution as a “March of Progress”». SAPIENS (en inglés estadounidense). Consultado el 11 de febrero de 2023.
- ↑ Strait, David; Grine, Frederick E.; Fleagle, John G. (2015). Henke, Winfried, ed. Analyzing Hominin Phylogeny: Cladistic Approach (en inglés). Springer Berlin Heidelberg. pp. 1989-2014. ISBN 978-3-642-39978-7. doi:10.1007/978-3-642-39979-4_58. Consultado el 17 de abril de 2025.
- ↑ Gunz, Philipp; Tilot, Amanda K.; Wittfeld, Katharina; Teumer, Alexander; Shapland, Chin Yang; van Erp, Theo G.M.; Dannemann, Michael; Vernot, Benjamin et al. (2019-01). «Neandertal Introgression Sheds Light on Modern Human Endocranial Globularity». Current Biology 29 (1): 120-127.e5. ISSN 0960-9822. doi:10.1016/j.cub.2018.10.065. Consultado el 17 de abril de 2025.
- ↑ Maslin, Mark A.; Shultz, Susanne; Trauth, Martin H. (5 de marzo de 2015). «A synthesis of the theories and concepts of early human evolution». Philosophical Transactions of the Royal Society B: Biological Sciences 370 (1663): 20140064. ISSN 0962-8436. doi:10.1098/rstb.2014.0064. Consultado el 17 de abril de 2025.
- ↑ a b Bastir, Markus; Rosas, Antonio; Gunz, Philipp; Peña-Melian, Angel; Manzi, Giorgio; Harvati, Katerina; Kruszynski, Robert; Stringer, Chris et al. (13 de diciembre de 2011). «Evolution of the base of the brain in highly encephalized human species». Nature Communications (en inglés) 2 (1). ISSN 2041-1723. doi:10.1038/ncomms1593. Consultado el 14 de abril de 2025.
- ↑ Holloway, Ralph L.; Sherwood, Chet C.; Hof, Patrick R.; Rilling, James K. (2009). Binder, Marc D., ed. Evolution of the Brain in Humans – Paleoneurology (en inglés). Springer. pp. 1326-1334. ISBN 978-3-540-29678-2. doi:10.1007/978-3-540-29678-2_3152. Consultado el 17 de abril de 2025.
- ↑ Gage, W. H., Winter, D. A., Frank, J. S., Adkin, A. L. (abril de 2004). «Kinematic and kinetic validity of the inverted pendulum model in quiet standing.»
- ↑ Aiello, Leslie C.; Wheeler, Peter (1995). «The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution». Current Anthropology (en inglés) 36 (2): 199-221. ISSN 0011-3204. doi:10.1086/204350. Consultado el 15 de abril de 2025.
- ↑ R. Leonard; Snodgrass; Robertson (2007). «Effects of Brain Evolution on Human Nutrition and Metabolism». Review in Advance (en inglés): 1-21. doi:10.1146/annurev.nutr.27.061406.093659. Consultado el 15 de abril de 2025.
- ↑ Ruff, Christopher B. (1994). «Morphological adaptation to climate in modern and fossil hominids». American Journal of Physical Anthropology (en inglés) 37 (S19): 65-107. ISSN 0002-9483. doi:10.1002/ajpa.1330370605. Consultado el 15 de abril de 2025.
- ↑ a b McHenry, Henry M. (1992). «Body size and proportions in early hominids». American Journal of Physical Anthropology (en inglés) 87 (4): 407-431. ISSN 0002-9483. doi:10.1002/ajpa.1330870404. Consultado el 15 de abril de 2025.
- ↑ a b Baab, Karen L.; McNulty, Kieran P.; Rohlf, F. James (2012). «The shape of human evolution: A geometric morphometrics perspective». Evolutionary Anthropology: Issues, News, and Reviews (en inglés) 21 (4): 151-165. ISSN 1060-1538. doi:10.1002/evan.21320. Consultado el 15 de abril de 2025.
- ↑ Ruff, Christopher (2002). «Variation in Human Body Size and Shape». Annual Review of Anthropology (en inglés) 31 (1): 211-232. ISSN 0084-6570. doi:10.1146/annurev.anthro.31.040402.085407. Consultado el 15 de abril de 2025.
- ↑ Gustafsson, Anders; Lindenfors, Patrik (2009-01). «Latitudinal patterns in human stature and sexual stature dimorphism». Annals of Human Biology (en inglés) 36 (1): 74-87. ISSN 0301-4460. doi:10.1080/03014460802570576. Consultado el 15 de abril de 2025.
- ↑ NCD Risk Factor Collaboration (NCD-RisC) (26 de julio de 2016). «A century of trends in adult human height». eLife (en inglés) 5. ISSN 2050-084X. PMC 4961475. PMID 27458798. doi:10.7554/eLife.13410. Consultado el 15 de abril de 2025.
- ↑ Bowler & Morus.
- ↑ Gowlett, J. A. J. (5 de junio de 2016). «The discovery of fire by humans: a long and convoluted process». Philosophical Transactions of the Royal Society B: Biological Sciences 371 (1696): 20150164. ISSN 0962-8436. doi:10.1098/rstb.2015.0164. Consultado el 14 de abril de 2025.
- ↑ Holloway, Ralph L.; Broadfield, Douglas C.; Yuan, Michael S. (6 de mayo de 2004). The Human Fossil Record. Wiley. ISBN 978-0-471-41823-8. Consultado el 14 de abril de 2025.
- ↑ Falk, Dean (1 de mayo de 2014). «Interpreting sulci on hominin endocasts: old hypotheses and new findings». Frontiers in Human Neuroscience 8. ISSN 1662-5161. doi:10.3389/fnhum.2014.00134. Consultado el 14 de abril de 2025.
- ↑ a b c Rosenberg, Karen; Trevathan, Wenda (2002). «Birth, obstetrics and human evolution». BJOG: An International Journal of Obstetrics & Gynaecology (en inglés) 109 (11): 1199-1206. ISSN 1470-0328. doi:10.1046/j.1471-0528.2002.00010.x. Consultado el 14 de abril de 2025.
- ↑ Tobias, Phillip V. (1987). «The brain of Homo habilis: A new level of organization in cerebral evolution». Journal of Human Evolution (en inglés) 16 (7-8): 741-761. doi:10.1016/0047-2484(87)90022-4. Consultado el 14 de abril de 2025.
- ↑ «Neotenia: el defecto que nos permitió tener un encéfalo más grande - Antroporama». Antroporama. 9 de abril de 2013. Archivado desde el original el 20 de octubre de 2021. Consultado el 13 de abril de 2025.
- ↑ Gómez Robles; C. Sherwood (8 de febrero de 2016). «La evolución del cerebro humano». La evolución del cerebro humano. doi:10.7203/metode.7.7602. Consultado el 13 de abril de 2025.
- ↑ Sutou, Shizuyo (8 de marzo de 2012). «Hairless mutation: a driving force of humanization from a human–ape common ancestor by enforcing upright walking while holding a baby with both hands». Genes to Cells 17 (4): 264-272. ISSN 1356-9597. doi:10.1111/j.1365-2443.2012.01592.x. Consultado el 10 de agosto de 2025.
- ↑ Ibraimov, A.I. (2007-02). «The Evolution of Body Heat Conductivity, Skin and Brain Size in Human». Journal of Human Ecology (en inglés) 21 (2): 95-103. ISSN 0970-9274. doi:10.1080/09709274.2007.11905956. Consultado el 10 de agosto de 2025.
- ↑ Lasisi, Tina; Smallcombe, James W.; Kenney, W. Larry; Shriver, Mark D.; Zydney, Benjamin; Jablonski, Nina G.; Havenith, George (13 de junio de 2023). «Human scalp hair as a thermoregulatory adaptation». Proceedings of the National Academy of Sciences 120 (24): e2301760120. PMC 10268308. PMID 37279270. doi:10.1073/pnas.2301760120. Consultado el 10 de agosto de 2025.
- ↑ Ruxton, Graeme D.; Wilkinson, David M. (27 de diciembre de 2011). «Avoidance of overheating and selection for both hair loss and bipedality in hominins». Proceedings of the National Academy of Sciences 108 (52): 20965-20969. PMC 3248486. PMID 22160694. doi:10.1073/pnas.1113915108. Consultado el 10 de agosto de 2025.
- ↑ a b c Rantala, M. J. (20 de agosto de 2007). «Evolution of nakedness in Homo sapiens». Journal of Zoology 273 (1): 1-7. ISSN 0952-8369. doi:10.1111/j.1469-7998.2007.00295.x. Consultado el 10 de agosto de 2025.
- ↑ Jablonski, N. G.; Chaplin, G. (2000-07). «The evolution of human skin coloration». Journal of Human Evolution 39 (1): 57-106. ISSN 0047-2484. PMID 10896812. doi:10.1006/jhev.2000.0403. Consultado el 10 de agosto de 2025.
- ↑ Quintero, Erika Montserrat Vilchis. «El mono desnudo: los humanos y la pérdida evolutiva del pelo corporal». ciencia.nexos.com.mx. Consultado el 10 de agosto de 2025.
- ↑ Las causas son debatidas, siendo el motivo principal del clásico El mono desnudo de Desmond Morris
- ↑ Véase El mono desnudo de Desmond Morris.
- ↑ Los seres humanos tienen una densidad capilar similar a la de los chimpancés, que es significativamente menor que la de los macacos. Por el contrario, la densidad de las glándulas ecrinas es, en promedio, 10 veces mayor en los seres humanos que en los chimpancés y los macacos, cuya densidad es sorprendentemente similar. J Hum Evol. 2018. Comparative evidence for the independent evolution of hair and sweat gland traits in primates.
- ↑ Kamberov, Yana G.; Guhan, Samantha M.; DeMarchis, Alessandra; Jiang, Judy; Wright, Sara Sherwood; Morgan, Bruce A.; Sabeti, Pardis C.; Tabin, Clifford J. et al. (2018-12). «Comparative evidence for the independent evolution of hair and sweat gland traits in primates». Journal of Human Evolution 125: 99-105. ISSN 1095-8606. PMC 6289065. PMID 30502901. doi:10.1016/j.jhevol.2018.10.008. Consultado el 10 de agosto de 2025.
- ↑ «Por qué los humanos, a diferencia de otros primates, no tenemos el cuerpo completamente cubierto de pelo». BBC News Mundo. Consultado el 10 de agosto de 2025.
- ↑ Giles, James (2010-12). «Naked Love: The Evolution of Human Hairlessness». Biological Theory 5 (4): 326-336. ISSN 1555-5542. doi:10.1162/biot_a_00062. Consultado el 10 de agosto de 2025.
- ↑ deMenocal, Peter B (2004-03). «African climate change and faunal evolution during the Pliocene–Pleistocene» [Cambio climático africano y evolución de la fauna durante el Plioceno-Pleistoceno]. Earth and Planetary Science Letters (en inglés) 220 (1-2): 3-24. doi:10.1016/S0012-821X(04)00003-2. Consultado el 13 de abril de 2025.
- ↑ Ungar, Peter S.; Grine, Frederick E.; Teaford, Mark F. (1 de octubre de 2006). «Diet in EarlyHomo: A Review of the Evidence and a New Model of Adaptive Versatility» [Dieta en los primeros Homo: Una revisión de la evidencia y un nuevo modelo de versatilidad adaptativa]. Annual Review of Anthropology 35 (1): 209-228. ISSN 0084-6570. doi:10.1146/annurev.anthro.35.081705.123153. Consultado el 13 de abril de 2025.
- ↑ Aiello, Leslie C.; Wheeler, Peter (1995). «The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution». Current Anthropology 36 (2): 199-221. ISSN 0011-3204. Consultado el 13 de abril de 2025.
- ↑ Wrangham, Richard W.; Jones, James Holland; Laden, Greg; Pilbeam, David; Conklin‐Brittain, NancyLou (1999-12). «The Raw and the Stolen» [Lo crudo y lo robado: La cocina y la ecología, orígenes humanos]. Current Anthropology 40 (5): 567-594. ISSN 0011-3204. doi:10.1086/300083. Consultado el 13 de abril de 2025.
- ↑ Marmelada, C.A. (2008). «Prolongación de la infancia y origen de la adolescencia en la evolución humana». Ciencia Cognitiva: Revista Electrónica de Divulgación. Consultado el 13 de abril de 2025.
- ↑ Leigh, S.R. (2004-03). «Brain growth, life history, and cognition in primate and human evolution». American Journal of Primatology (en inglés) 62 (3): 139-164. ISSN 0275-2565. doi:10.1002/ajp.20012. Consultado el 13 de abril de 2025.
- ↑ Terrence Deacon, The Symbolic Species. Fuente citada en en:The Symbolic Species.
- ↑ "La clave del origen del lenguaje está en los huesos del oído hallados en Atapuerca" - Ignacio Mendizábal explica en Burgos que el origen de la expresión oral hay que buscarlo más allá de un millón de años, DCYT, 14 de diciembre de 2010.
- ↑ a b Pausas, Juli G.; Keeley, Jon E. (1 de julio de 2009). «A Burning Story: The Role of Fire in the History of Life». BioScience 59 (7): 593-601. ISSN 0006-3568. doi:10.1525/bio.2009.59.7.10. Consultado el 9 de agosto de 2025.
- ↑ "En la mayor parte del Viejo Mundo —África, Eurasia—, el fuego antropogénico avanzó poco a poco. La antorcha pasó de un homínido a otro; los paisajes ya adaptados a un régimen de fuego provocado por el hombre aceptaron otro". Sthephen J. Pyne (2019). Fire: A Brief History. NewSouth Publishing.
- ↑ a b c Pyne, Stephen (2019). Fire: A Brief History (en inglés.). Australia: NewSouth Publishing. ISBN 9781742249391. Consultado el 11-08-2025.
- ↑ a b He, Tianhua; Lamont, Byron B.; Pausas, Juli G. (2019). «Fire as a key driver of Earth's biodiversity». Biological Reviews (en inglés) 94 (6): 1983-2010. ISSN 1469-185X. doi:10.1111/brv.12544. Consultado el 9 de agosto de 2025.
- ↑ Eiseley, Loren C. (1954). «Man the Fire-Maker». Scientific American 191 (3): 52-57. ISSN 0036-8733. Consultado el 9 de agosto de 2025.
- ↑ Berna, Francesco; Goldberg, Paul; Horwitz, Liora Kolska; Brink, James; Holt, Sharon; Bamford, Marion; Chazan, Michael (15 de mayo de 2012). «Microstratigraphic evidence of in situ fire in the Acheulean strata of Wonderwerk Cave, Northern Cape province, South Africa». Proceedings of the National Academy of Sciences 109 (20): E1215-E1220. PMC 3356665. PMID 22474385. doi:10.1073/pnas.1117620109. Consultado el 9 de agosto de 2025.
- ↑ a b Wrangham, Richard; Carmody, Rachel (2010). «Human Adaptation to the Control of Fire». Wiley-Blackwell (en inglés estadounidense). ISSN 1060-1538. Consultado el 10 de agosto de 2025.
- ↑ Carbonell, Eudald (2024). «¿Qué hemos comido?». De la caverna al cosmos. Nuestro futuro como especie. Barcelona: RBA. p. 194. ISBN 978-8411730969.
- ↑ Zohar, Irit; Alperson-Afil, Nira; Goren-Inbar, Naama; Prévost, Marion; Tütken, Thomas; Sisma-Ventura, Guy; Hershkovitz, Israel; Najorka, Jens (2022-12). «Evidence for the cooking of fish 780,000 years ago at Gesher Benot Ya’aqov, Israel». Nature Ecology & Evolution (en inglés) 6 (12): 2016-2028. ISSN 2397-334X. doi:10.1038/s41559-022-01910-z. Consultado el 9 de agosto de 2025.
- ↑ Goren-Inbar, Naama (2017). Bar-Yosef, Ofer, ed. Gesher Benot Ya‘aqov. Cambridge University Press. pp. 187-194. ISBN 978-1-107-09046-0. Consultado el 9 de agosto de 2025.
- ↑ Roebroeks, Wil; Villa, Paola (29 de marzo de 2011). «On the earliest evidence for habitual use of fire in Europe». Proceedings of the National Academy of Sciences (en inglés) 108 (13): 5209-5214. ISSN 0027-8424. doi:10.1073/pnas.1018116108. Consultado el 11 de agosto de 2025.
- ↑ «Ways of Catching a Spark: A History of Fire-Making Methods | Science Museum». Science Museum (en inglés). Archivado desde el original el 13 de agosto de 2024. Consultado el 11 de agosto de 2025.
- ↑ Glikson, Andrew (1 de noviembre de 2013). «Fire and human evolution: The deep-time blueprints of the Anthropocene». Anthropocene 3: 89-92. ISSN 2213-3054. doi:10.1016/j.ancene.2014.02.002. Consultado el 9 de agosto de 2025.
- ↑ "La mejora de la digestibilidad como resultado de la cocción es del 12-35 % para los almidones (mediana = 30 %: avena, trigo, plátano macho, patata y plátano verde) y del 45-78 % para las proteínas (huevo de gallina). El coste energético de la cocción de alimentos actualmente no se mide, pero tendría que ser muy alto para anular estos beneficios". Wrangham; Carmody (2010). Human Adaptation to the Control of Fire
- ↑ K. Austin. «Fire and the Brain: How Cooking Shaped Humans». American Museum of Natural History. Consultado el 11 de agosto de 2025.
- ↑ "La asombrosa dispersión de los humanos por el planeta coincide con una ola de extinciones de la megafauna que implica al fuego como catalizador. Lo más probable es que el fuego contribuyera a esas extinciones". Sthephen J. Pyne (2019). Fire: A Brief History. NewSouth Publishing. p.31
- ↑ Chapais, Bernard (2013-03). «Monogamy, strongly bonded groups, and the evolution of human social structure». Evolutionary Anthropology: Issues, News, and Reviews (en inglés) 22 (2): 52-65. ISSN 1060-1538. doi:10.1002/evan.21345. Consultado el 16 de abril de 2025.
- ↑ Opie, Christopher; Atkinson, Quentin D.; Dunbar, Robin I. M.; Shultz, Susanne (13 de agosto de 2013). «Male infanticide leads to social monogamy in primates». Proceedings of the National Academy of Sciences (en inglés) 110 (33): 13328-13332. ISSN 0027-8424. doi:10.1073/pnas.1307903110. Consultado el 16 de abril de 2025.
- ↑ Lukas, D.; Clutton-Brock, T. H. (2 de agosto de 2013). «The Evolution of Social Monogamy in Mammals». Science (en inglés) 341 (6145): 526-530. ISSN 0036-8075. doi:10.1126/science.1238677. Consultado el 16 de abril de 2025.
| Control de autoridades |
|
|---|
 Datos: Q1362274
Datos: Q1362274