Respuesta de evitación de interferencias
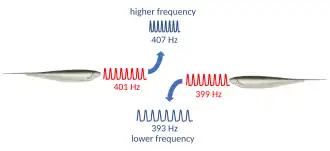
La respuesta de evitación de interferencias (JAR por sus siglas en inglés) es un comportamiento de algunas especies de peces débilmente eléctricos. Ocurre cuando dos peces eléctricos con descargas ondulatorias se encuentran: si sus frecuencias de descarga son muy similares, cada pez desplaza su frecuencia de descarga para aumentar la diferencia entre ambas. De este modo, ambos peces evitan la interferencia de su sentido de electrorrecepción.
El comportamiento se ha estudiado más intensamente en la especie sudamericana Eigenmannia virescens. También está presente en otros Gymnotiformes como Apteronotus, así como en la especie africana Gymnarchus niloticus. La respuesta de evitación de interferencias fue una de las primeras respuestas conductuales complejas en un vertebrado en tener su circuito neural completamente especificado. Como tal, reviste especial importancia en el campo de la neuroetología.
Descubrimiento
La respuesta de evitación de interferencia (JAR) fue descubierta por Akira Watanabe y Kimihisa Takeda en 1963. El pez que utilizaron era una especie no especificada de Eigenmannia, que tiene una descarga de onda casi sinusoidal de unos 300 Hz. Descubrieron que cuando se emite un estímulo eléctrico sinusoidal desde un electrodo cerca del pez, si la frecuencia del estímulo está dentro de los 5 Hz de la frecuencia de descarga eléctrica del órgano (EOD) del pez, éste altera su frecuencia EOD para aumentar la diferencia entre su propia frecuencia y la frecuencia del estímulo. Los estímulos por encima de la frecuencia EOD del pez empujan la frecuencia EOD hacia abajo, mientras que las frecuencias por debajo de la del pez empujan la frecuencia EOD hacia arriba, con un cambio máximo de unos ±6,5 Hz.[1] Este comportamiento recibió el nombre de «respuesta de evitación de interferencias» varios años después, en 1972, en un artículo de Theodore Bullock, Robert Hamstra Jr. y Henning Scheich.[2]

En 1975, Walter Heiligenberg descubrió un JAR en Gymnarchus niloticus, demostrando que el comportamiento había evolucionado de forma convergente en dos linajes separados.[3]
Comportamiento
Eigenmannia y otros peces débilmente eléctricos utilizan la electrorrecepción activa: pueden localizar objetos generando un campo eléctrico y detectando distorsiones en el campo causadas por interferencias de esos objetos. Los peces eléctricos utilizan su órgano eléctrico para crear campos eléctricos y detectan pequeñas distorsiones de estos campos mediante órganos electrorreceptores especiales en la piel. Todos los peces con JAR son peces con descargas ondulatorias que emiten descargas cuasi-sinusoidales estables. Para el género Eigenmannia, las frecuencias oscilan entre 240 y 600 Hz.[4] La frecuencia de la EOD es muy constante, normalmente con una variación inferior al 0,3% en un intervalo de tiempo de 10 minutos.[2]
Si un campo eléctrico sinusoidal vecino está descargando cerca de la frecuencia EOD del pez, causa interferencias que provocan confusión sensorial en el pez e interferencias suficientes para impedirle una electrolocalización eficaz.[5] Eigenmannia suelen estar dentro del rango de campo eléctrico de tres a cinco peces de la misma especie en cualquier momento. Si muchos peces se encuentran cerca unos de otros, es beneficioso para cada pez distinguir entre su propia señal y las de los demás; esto puede hacerse aumentando la diferencia de frecuencia entre sus descargas. Por lo tanto, parece que la función del JAR es evitar la confusión sensorial entre peces vecinos.[6]
Para determinar lo cerca que está la frecuencia del estímulo de la frecuencia de descarga, el pez compara las dos frecuencias utilizando sus órganos electrorreceptores, en lugar de comparar la frecuencia de descarga con un marcapasos interno; en otras palabras, el JAR se basa únicamente en información sensorial. Esto se determinó experimentalmente silenciando el órgano eléctrico de un pez con curare y estimulándolo después con dos frecuencias externas. El JAR, medido a partir de las neuronas electromotoras de la médula espinal, dependía únicamente de las frecuencias de los estímulos externos, y no de la frecuencia del marcapasos.[7]
Neurobiología
Eigenmannia (Gymnotiformes)
La mayor parte de la vía JAR en los Gymnotiformes sudamericanos se ha elaborado utilizando Eigenmannia virescens como sistema modelo.[8][9]
Codificación sensorial

Cuando la frecuencia del estímulo y la frecuencia de descarga están próximas entre sí, las dos ondas de amplitud-tiempo sufren interferencias, y los órganos electrorreceptivos perciben una única onda con una frecuencia intermedia. Además, la onda combinada estímulo-EOD tiene un patrón rítmico, con la frecuencia de batido igual a la diferencia de frecuencia entre el estímulo y el EOD.[10]
Los gimnotiformes tienen dos clases de órganos electrorreceptores, los receptores ampulares y los receptores tuberosos. Los receptores ampulares responden a estímulos de baja frecuencia, inferiores a 40 Hz, y actualmente se desconoce su función en el JAR. Los receptores tuberosos responden a frecuencias más altas, disparando mejor cerca de la frecuencia normal de EOD del pez. Los receptores tuberosos propiamente dichos son de dos tipos, la unidad T y la unidad P. La unidad T (T significa tiempo, es decir, fase en el ciclo) se dispara de forma sincrónica con la frecuencia de la señal disparando un pico en cada ciclo de la forma de onda. Las unidades P (P significa probabilidad) tienden a disparar cuando la amplitud aumenta y disparan menos cuando disminuye. En condiciones de interferencia, la unidad P se dispara en los picos de amplitud del ciclo de latido, donde las dos ondas interfieren constructivamente. Por lo tanto, una señal combinada de estímulo y EOD hace que las unidades T se disparen en la frecuencia intermedia y que las unidades P aumenten y disminuyan periódicamente con el latido.[7]
Procesamiento en el cerebro
Las unidades T de codificación temporal convergen en neuronas llamadas células esféricas en el lóbulo de la línea lateral electrosensorial. Al combinar la información de varias unidades T, la célula esférica es aún más precisa en su codificación temporal. Las unidades P que codifican la amplitud convergen en células piramidales, también en el lóbulo de la línea lateral electrosensorial. Existen dos tipos de células piramidales: las unidades E excitatorias, que disparan más cuando son estimuladas por las unidades P, y las unidades I inhibitorias, que disparan menos cuando son estimuladas por interneuronas inhibitorias activadas por las unidades P.[8]
A continuación, las células esféricas y las células piramidales se proyectan al torus semicircularis, una estructura con muchas láminas (capas) en el mesencéfalo. Aquí se integra la información de fase y amplitud para determinar si la frecuencia del estímulo es mayor o menor que la frecuencia EOD. Las neuronas signo-selectivas de las capas más profundas del torus semicircularis son selectivas a si la diferencia de frecuencia es positiva o negativa; cualquier célula signo-selectiva se dispara en un caso pero no en el otro.[9]
Salida
Las células selectivas de señales entran en el núcleo electrosensorius del diencéfalo, que a su vez se proyecta en dos vías diferentes. Las neuronas selectivas para una diferencia positiva (estímulo superior al EOD) estimulan el núcleo prepacemaker, mientras que las neuronas selectivas para una diferencia negativa (estímulo inferior al EOD) inhiben el núcleo prepacemaker sublemniscal. Ambos núcleos prepacemaker envían proyecciones al núcleo pacemaker, que en última instancia controla la frecuencia del EOD.[8]
Gymnarchus (Osteoglossiformes)
La vía neural del JAR en Gymnarchus es casi idéntica a la de los Gymnotiformes, con algunas pequeñas diferencias. Las unidades S de Gymnarchus codifican el tiempo, como las unidades T de Gymnotiformes. Las unidades O codifican la intensidad de la señal, como las unidades P de los Gymnotiformes, pero responden en un rango más estrecho de intensidades. En Gymnarchus, las diferencias de fase entre el EOD y el estímulo se calculan en el lóbulo de la línea lateral electrosensorial y no en el torus semicircularis.[11]
Filogenia y evolución de los peces débilmente eléctricos
Existen dos órdenes principales de peces débilmente eléctricos, los Gymnotiformes de Sudamérica y los Osteoglossiformes de África. Lo más probable es que la electrorrecepción surgiera de forma independiente en los dos linajes. Los peces débilmente eléctricos son en su mayoría descargadores de impulsos, que no realizan el JAR, mientras que algunos son descargadores de ondas. La descarga de ondas evolucionó en dos taxones: la superfamilia Apteronotoidea (orden Gymnotiformes) y la especie Gymnarchus niloticus (orden Osteoglossiformes). Entre los géneros notables de Apteronotoidea que realizan JAR se incluyen Eigenmannia y Apteronotus.[12] Aunque evolucionaron el JAR por separado, los taxones sudamericanos y africanos (en negrita en el árbol) han evolucionado de forma convergente mecanismos computacionales neurales y respuestas conductuales casi idénticos para evitar la interferencia, con sólo pequeñas diferencias.[13] La filogenia de los clados de peces débilmente eléctricos, omitiendo los peces no eléctricos y fuertemente eléctricos, muestra los principales acontecimientos en su evolución.[12][14] En el árbol, «sp» significa «una especie» y “spp” significa «múltiples especies».
| Vertebrados |
| ||||||||||||||||||||||||||||||||||||

Véase también
Referencias
- ↑ Watanabe, Akira; Takeda, Kimihisa (1963). «"The change of discharge frequency by A.C. stimulus in a weak electric fish"». The Journal of Experimental Biology. doi:10.1242/jeb.40.1.57.
- ↑ a b Bullock, Theodore H.; Hamstra, R. Jr.; Scheich, H. (1972). «"The jamming avoidance response of high frequency electric fish".». Journal of Comparative Physiology. doi:10.1007/BF00696517.
- ↑ Heiligenberg, Walter (1975). «"Electrolocation and jamming avoidance in the electric fish Gymnarchus niloticus (Gymnarchidae, Mormyriformes)"». Journal of Comparative Physiology A. doi:10.1007/bf01380044.
- ↑ Hopkins, C. (1974). «"Electric communication: functions in the social behavior of Eigenmannia virescens"». Behaviour. doi:10.1163/156853974X00499.
- ↑ Shifman, Aaron R.; Lewis, John E. (2018-01). «The complexity of high-frequency electric fields degrades electrosensory inputs: implications for the jamming avoidance response in weakly electric fish». Journal of the Royal Society, Interface 15 (138): 20170633. ISSN 1742-5662. PMC 5805966. PMID 29367237. doi:10.1098/rsif.2017.0633. Consultado el 4 de junio de 2025.
- ↑ Tan, E.; Nizar, J.; Carrera-G, E.; Fortune, E. (2005). «"Electrosensory interference in naturally occurring aggregates of a species of weakly electric fish, Eigenmannia virescens"». Behavioural Brain Research. PMID 16099058. doi:10.1016/j.bbr.2005.06.014.
- ↑ a b Scheich, H.; Bullock, Theodore H.; Hamstra, R. Jr. (1973). «"Coding properties of two classes of afferent nerve fibers: high-frequency electroreceptors in the electric fish, Eigenmannia".». Journal of Neurophysiology. PMID 4705666. doi:10.1152/jn.1973.36.1.39.
- ↑ a b c Bastian, J.; Heiligenberg, Walter (1980). «"Neural correlates of the jamming avoidance response of Eigenmannia".». Journal of Comparative Physiology A. doi:10.1007/BF00656908.
- ↑ a b Heiligenberg, W.; Rose, G. (1985-02). «Phase and amplitude computations in the midbrain of an electric fish: intracellular studies of neurons participating in the jamming avoidance response of Eigenmannia». The Journal of Neuroscience: The Official Journal of the Society for Neuroscience 5 (2): 515-531. ISSN 0270-6474. PMC 6565201. PMID 3973680. doi:10.1523/JNEUROSCI.05-02-00515.1985. Consultado el 5 de junio de 2025.
- ↑ Kramer, Berndt (1999). «"Waveform discrimination, phase sensitivity and jamming avoidance in a wave-type electric fish".». Journal of Experimental Biology. PMID 10210679. doi:10.1242/jeb.202.10.1387.
- ↑ Bullock, T.; Behrend, K.; Heiligenberg, Walter (1975). «"Comparison of the jamming avoidance responses in Gymnotid and Gymnarchid electric fish: A case of convergent evolution of behavior and its sensory basis".». Journal of Comparative Physiology. doi:10.1007/BF01380047.
- ↑ a b Bullock, T. H.; Bodznick, D. A.; Northcutt, R. G. (1983). «"The phylogenetic distribution of electroreception: Evidence for convergent evolution of a primitive vertebrate sense modality"». Brain Research Reviews. PMID 6616267. doi:10.1016/0165-0173(83)90003-6.
- ↑ Kawasaki, M. (1975). «"Independently evolved jamming avoidance responses in Gymnotid and Gymnarchid electric fish: a case of convergent evolution of behavior and its sensory basis".». Journal of Comparative Physiology. PMID 8366474. doi:10.1007/BF00209614.
- ↑ Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; Sullivan, John P.; Hopkins, Carl D.; Nishida, Mutsumi (14 de mayo de 2012). «Comparable Ages for the Independent Origins of Electrogenesis in African and South American Weakly Electric Fishes». En Murphy, William J., ed. PLoS ONE (en inglés) 7 (5): e36287. ISSN 1932-6203. PMC 3351409. PMID 22606250. doi:10.1371/journal.pone.0036287. Consultado el 5 de junio de 2025.
Lectura adicional
- Heiligenberg, W. (1977) Principles of Electrolocation and Jamming Avoidance in Electric Fish: A Neuroethological Approach. Studies of Brain Function, Vol. 1. Berlin-New York: Springer Verlag.
- Heiligenberg, W. (1990) Electric Systems in Fish. Synapse 6:196-206.
- Heiligenberg, W. (1991) Neural Nets in Electric fish. MIT Press: Cambridge, Massachusetts.
- Kawasaki, M. (2009) Evolution of time-coding systems in weakly electric fishes. Zoological Science 26: 587-599.
| Control de autoridades |
|
|---|
 Datos: Q6147980
Datos: Q6147980