Evolución del veneno de serpiente
_juvenile_eating_(14232058164).jpg)
El veneno en serpientes y algunos lagartos es una forma de saliva que ha sido modificada en veneno a lo largo de su historia evolutiva.[1] En las serpientes, el veneno ha evolucionado para matar o someter a la presa, así como para realizar otras funciones relacionadas con la dieta.[2] Aunque las serpientes ocasionalmente usan su veneno en defensa propia, no se cree que esto haya tenido un efecto significativo en la evolución del veneno.[3] Se considera que la evolución del mismo es responsable de la enorme expansión de las serpientes por todo el mundo.[4][5][6]
La historia evolutiva del veneno de serpiente es un tema de debate. Históricamente, se creía que el veneno de serpiente había evolucionado una sola vez, en la base de las Caenophidia, o serpientes derivadas. Estudios moleculares publicados a partir de 2006 sugirieron que el veneno se originó una sola vez en un supuesto clado de reptiless, llamado Toxicofera, hace aproximadamente 170 millones de años.[7] Bajo esta hipótesis, el veneno toxicoferano original era un conjunto muy simple de proteínas que se ensamblaban en un par de glándulas. Posteriormente, este conjunto de proteínas se diversificó en las diversas líneas de toxicoferanos, incluyendo Serpentes, Anguimorpha e Iguania: varias líneas de serpientes también perdieron la capacidad de producir veneno.[8][9] La hipótesis toxicoferana fue cuestionada por estudios a mediados de la década de 2010, incluido un estudio de 2015 que encontró que las proteínas del veneno tenían homólogos en muchos otros tejidos en la pitón birmana.[10][11] El estudio sugirió, por lo tanto, que el veneno había evolucionado de manera independiente en diferentes linajes de reptiles, incluyendo una vez en las serpientes caenofidias.[10] Se cree que el veneno que contiene la mayoría de las familias de toxinas existentes estaba presente en el ancestro común más reciente de las Caenophidia: estas toxinas posteriormente experimentaron una enorme diversificación, acompañada de cambios en la morfología de las glándulas de veneno y los sistemas de entrega.[12]
Se cree que la evolución del veneno de serpiente está impulsada por una carrera armamentística evolutiva entre las proteínas del veneno y la fisiología de la presa.[13] El mecanismo común de evolución se considera la duplicación génica seguida de selección natural para rasgos adaptativos.[14] Las adaptaciones producidas por este proceso incluyen un veneno más tóxico para presas específicas en varias líneas,[15][16][17] proteínas que pre-digieren a la presa,[18] y un método para rastrear a la presa después de una mordedura.[19] Estas diversas adaptaciones del veneno también han llevado a un considerable debate sobre la definición de veneno y serpientes venenosas.[20] Los cambios en la dieta de un linaje se han relacionado con la atrofia del veneno.[8][9]
Historia evolutiva
Se cree que el origen del veneno proporcionó el catalizador para la rápida diversificación de las serpientes en el período Cenozoico,[21] particularmente para las Colubridae y su colonización de las Américas.[5] Los estudiosos sugieren que la razón de esta gran expansión fue el cambio de un método mecánico a un método bioquímico para someter a la presa.[22][23] Los venenos de serpiente atacan vías y procesos biológicos que también son objetivo de los venenos de otros taxones; por ejemplo, se han encontrado bloqueadores de canal de calcio en serpientes, arañas y caracoles cono, lo que sugiere que el veneno exhibe evolución convergente.[24] El veneno es común entre las familias de serpientes derivadas.[13] Se cree que el veneno que contiene la mayoría de las familias de toxinas existentes estaba presente en el ancestro común más reciente de las Caenophidia, también llamadas Colubroidea. Estas toxinas posteriormente experimentaron una tremenda diversificación, acompañada de cambios en la morfología de las glándulas de veneno y los sistemas de entrega. Esta diversificación está vinculada a la rápida radiación global de las serpientes avanzadas.[12] Los colmillos tubulares o acanalados que las serpientes usan para administrar su veneno a su objetivo han evolucionado varias veces y son un ejemplo de evolución convergente. Los colmillos tubulares comunes a las serpientes con colmillos frontales se cree que evolucionaron de manera independiente en Viperidae, Elapidae y Atractaspidinae.[25]
| Serpientes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
.jpg)
Un cladograma adaptado de Fry et al. (2012) que muestra un subconjunto de eventos sugeridos de reclutamiento de proteínas. [A]: 13 familias de toxinas, incluyendo 3FTx y metaloproteasa. [B]: 2 familias de toxinas, incluyendo PLA2 Tipo IIA y la clase P-II de metaloproteasas de veneno de serpiente. [C]: 2 familias de toxinas, incluyendo PLA2 Tipo IB.[26][27]
Hasta que el uso de la secuenciación genética para crear árboles filogenéticos se volvió práctico, las filogenias se creaban en base a la morfología. Dichas filogenias tradicionales sugerían que el veneno se originó a lo largo de múltiples ramas entre los Squamata hace aproximadamente 100 millones de años: en las Caenophidia, o serpientes derivadas, y en el género de lagartos Heloderma.[7][10][28] Estudios que utilizaron secuencias de gen nuclear en la década de 2000 y principios de 2010 encontraron la presencia de proteínas de veneno en los clados de lagartos Anguimorpha e Iguania similares a las de las serpientes, y sugirieron que, junto con Serpentes, estos formaban un clado, al que nombraron "Toxicofera".[7][28] Esto llevó a la teoría de que el veneno se originó solo una vez dentro de todo el linaje hace aproximadamente 170 millones de años. Este veneno ancestral se describió como un conjunto muy simple de proteínas, ensambladas en un par de glándulas.[7] Los venenos de los diferentes linajes luego se diversificaron y evolucionaron de manera independiente, junto con sus medios para inyectar veneno en la presa.[7] Esta diversificación incluyó la evolución independiente del sistema de entrega de veneno con colmillos frontales desde el sistema ancestral de entrega de veneno con colmillos traseros.[29] La hipótesis de origen único también sugiere que los sistemas de veneno posteriormente se atrofiaron, o se perdieron completamente, de manera independiente en varios linajes.[20] La posición filogenética de Iguania dentro de Toxicofera está respaldada por la mayoría de los estudios moleculares, pero no por los morfológicos.[30]
La hipótesis toxicoferana fue posteriormente cuestionada. Un estudio realizado en 2014 encontró que los homólogos de 16 proteínas de veneno, que se habían utilizado para respaldar la hipótesis de origen único, estaban todos expresados en altos niveles en varios tejidos del cuerpo.[11] Por lo tanto, los autores sugirieron que investigaciones previas, que habían encontrado proteínas de veneno conservadas a través del supuesto linaje toxicoferano, podrían haber malinterpretado la presencia de genes más genéricos de "mantenimiento" a través de este linaje, como resultado de solo muestrear ciertos tejidos dentro de los cuerpos de los reptiles. Por lo tanto, los autores propusieron que, en lugar de evolucionar solo una vez en un reptil ancestral, el veneno evolucionó de manera independiente en múltiples linajes, incluyendo una vez antes de la radiación de las serpientes "avanzadas".[11] Un estudio de 2015 encontró que los homólogos de los llamados genes "tóxicos" estaban presentes en numerosos tejidos de una serpiente no venenosa, la pitón birmana. Uno de los autores afirmó que habían encontrado homólogos de los genes de veneno en muchos tejidos fuera de las glándulas orales, donde se podrían esperar los genes de veneno. Esto demostró las debilidades de solo analizar transcriptomas (el total de ARN mensajero en una célula).[10] El equipo sugirió que las pitones representaban un período en la evolución de las serpientes antes del desarrollo importante del veneno. Los investigadores también encontraron que la expansión de las familias de genes de veneno ocurrió principalmente en las serpientes caenofidianas colubridas, lo que sugiere que la mayor parte de la evolución del veneno tuvo lugar después de que este linaje divergiera de otras serpientes.[10] El debate sobre la hipótesis toxicoferana está impulsado en parte por el desacuerdo sobre la definición de un veneno.[10][31] Hasta 2022, la hipótesis toxicoferana sigue siendo una visión predominante.[32]
Mecanismos de evolución
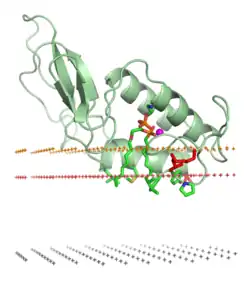
El mecanismo principal para la diversificación del veneno se cree que es la duplicación de genes que codifican para otros tejidos, seguida de su expresión en las glándulas de veneno. Las proteínas luego evolucionaron en varias proteínas de veneno a través de la selección natural. Este proceso, conocido como el modelo de nacimiento y muerte, es responsable de varios de los eventos de reclutamiento de proteínas en el veneno de serpiente.[33][13] Estas duplicaciones ocurrieron en una variedad de tipos de tejidos con varias funciones ancestrales. Ejemplos notables incluyen 3FTx, originalmente un neurotransmisor encontrado en el cerebro, que ha evolucionado en una neurotoxina que se une y bloquea los receptores de acetilcolina. Otro ejemplo es la fosfolipasa A2 (PLA2) tipo IIA, originalmente involucrada en procesos inflamatorios en tejidos normales, que ha evolucionado en un veneno capaz de desencadenar la actividad de la lipasa y la destrucción de tejidos.[21] El cambio en la función de PLA2, en particular, ha sido bien documentado; hay evidencia de varios eventos de duplicación génica separados, a menudo asociados con el origen de nuevas especies de serpientes.[34] La recombinación homóloga no alélica inducida por la invasión de transposones (o recombinación entre secuencias de ADN que son similares, pero no alelos) se ha propuesto como el mecanismo de duplicación de los genes PLA2 en las serpientes de cascabel, como una explicación para su rápida evolución.[35] Estas proteínas de veneno también han sido ocasionalmente reclutadas de nuevo en genes de tejidos.[36]
La duplicación génica no es la única forma en que el veneno se ha diversificado. Ha habido casos de nuevas proteínas de veneno generadas por empalme alternativo. La serpiente elápida Bungarus fasciatus, por ejemplo, posee un gen que se empalma alternativamente para producir tanto un componente de veneno como una proteína fisiológica.[36] La diversificación adicional puede haber ocurrido por la pérdida de genes de componentes específicos de veneno. Por ejemplo, se cree que el ancestro de las serpientes de cascabel tenía los genes PLA2 para una neurotoxina heterodimérica que ahora se encuentra en Crotalus scutulatus, pero esos genes están ausentes en las especies modernas de Crotalus no neurotóxicas; los genes PLA2 para la miotoxina Lys49, que supuestamente existía en el ancestro común de las serpientes de cascabel, también se perdieron varias veces en los linajes recientes hasta las especies existentes.[35] La pérdida de dominios también se ha implicado en la neofuncionalización del veneno. La investigación de la historia evolutiva de los genes de veneno SVMP de víboras reveló casos repetidos de pérdida de dominios, junto con una selección positiva significativa en la mayoría de las ramas filogenéticas donde se pensaba que había ocurrido la pérdida de dominios.[37] Los toxinas de veneno también han evolucionado a través del "secuestro" o "cooptación" de genes, o el cambio en la función de genes no relacionados. Un estudio de 2021 sugirió que la cooptación explicaba la evolución de la mayoría de los tipos de toxinas, pero no la de las toxinas más abundantes en el veneno de serpiente.[27]
Los eventos de reclutamiento de proteínas han ocurrido en diferentes puntos de la historia evolutiva de las serpientes. Por ejemplo, la familia de proteínas 3FTX está ausente en el linaje de víboras, lo que sugiere que fue reclutada en el veneno de serpiente después de que las víboras se separaran de las colubroides restantes.[38] Se cree que la PLA2 ha sido reclutada al menos dos veces por separado en el veneno de serpiente, una en los elápidos y otra en los víboras, mostrando una evolución convergente de esta proteína en una toxina.[39][40] Un estudio de 2019 sugirió que la duplicación génica podría haber permitido que diferentes toxinas evolucionaran de manera independiente, permitiendo a las serpientes experimentar con sus perfiles de veneno y explorar nuevas formulaciones de veneno efectivas.[40] Esto se propuso como una de las formas en que las serpientes han diversificado sus formulaciones de veneno a través de millones de años de evolución.[40] Los diversos eventos de reclutamiento han resultado en que el veneno de serpiente evolucione en una mezcla muy compleja de proteínas. El veneno de las serpientes de cascabel, por ejemplo, incluye cerca de 40 proteínas diferentes de diferentes familias de proteínas,[41] y se ha encontrado que otros venenos de serpiente contienen más de 100 proteínas distintas.[22] Se ha demostrado que la composición de esta mezcla varía geográficamente y está relacionada con las especies de presas disponibles en una región determinada.[17] El veneno de serpiente generalmente ha evolucionado muy rápidamente, con cambios que ocurren más rápido en el veneno que en el resto del organismo.[42]
Presión de selección
Las hipótesis de larga data sobre la evolución del veneno de serpiente han asumido que la mayoría de las serpientes inyectan mucho más veneno en su presa de lo necesario para matarla; por lo tanto, la composición del veneno no estaría sujeta a la selección natural. Esto se conoce como la hipótesis del "exceso de matanza".[43] Sin embargo, estudios recientes de la historia molecular del veneno de serpiente han contradicho esto, encontrando en cambio evidencia de una rápida evolución adaptativa en muchos clados diferentes, incluyendo las víboras alfombra, Echis,[44] las serpientes de cascabel de tierra, Sistrurus,[41] y la víbora malaya,[45] así como en general en la diversificación de las proteínas PLA2.[34] Hay evidencia filogenética de selección positiva y tasas rápidas de ganancia y pérdida de genes en los genes de veneno de los taxones Sistrurus que se alimentan de diferentes presas.[46] Hasta 2019, existía evidencia tanto de "exceso de matanza" en algunos linajes, como de una rápida evolución adaptativa y una carrera armamentística evolutiva con la fisiología de la presa en muchos otros.[47]
Los genes que codifican las proteínas de veneno en algunos géneros de serpientes tienen una proporción de mutaciones sinónimas menor a la esperada si el veneno estuviera evolucionando a través de procesos evolutivos neutrales; sin embargo, la tasa de mutaciones no sinónimas fue encontrada más alta en muchos casos, indicando una selección direccional.[34] Además, el veneno de serpiente es metabólicamente costoso para que una serpiente lo produzca, lo que los científicos han sugerido como evidencia adicional de que existe una presión de selección sobre el veneno de serpiente (en este caso, para minimizar el volumen de veneno requerido).[43] El uso de organismo modelo, en lugar de las presas naturales de las serpientes, para estudiar la toxicidad de la presa, se ha sugerido como una razón por la cual la hipótesis del "exceso de matanza" puede haber sido sobreestimada.[48] Sin embargo, el género de víboras de foseta Agkistrodon se ha encontrado como una excepción a esto; la composición del veneno en Agkistrodon se ha encontrado relacionada con la posición de la especie dentro de la filogenia, sugiriendo que esos venenos han evolucionado principalmente a través de procesos neutrales (mutación y deriva genética), y que puede haber una variación significativa en la presión de selección sobre varios venenos de serpiente.[49]
Varios estudios han encontrado evidencia de que el veneno y la resistencia al veneno en especies de presas han evolucionado en una coevolución armamentística.[43] Por ejemplo, las ratas de madera del género Neotoma tienen un alto grado de resistencia al veneno de las serpientes de cascabel, lo que sugiere que las ratas han evolucionado en respuesta al veneno de la serpiente, renovando así la presión de selección sobre las serpientes.[41] Se ha encontrado resistencia a los venenos de especies de serpientes depredadoras simpátricas en anguilas,[50] ardillas terrestres,[51] ardillas roqueras,[52] y ardillas grises del este.[53] Todos estos estudios sugirieron una carrera armamentística coevolutiva entre presa y depredador, indicando otra posible presión de selección sobre el veneno de serpiente para aumentar o innovar en toxicidad. La presión de selección sobre el veneno de serpiente se cree que selecciona la diversidad funcional dentro de las proteínas en el veneno, tanto dentro de una especie dada como entre especies.[54] Además de la fisiología de la presa, existe evidencia de que el veneno de serpiente ha evolucionado en respuesta a la fisiología de los depredadores.[13]
Además de la dieta, hay otras posibles presiones sobre la composición del veneno de serpiente. Un estudio de 2019 encontró que una mayor masa corporal y hábitats ecológicos más pequeños estaban correlacionados con un mayor rendimiento de veneno.[47] Otro estudio encontró que el clima y la temperatura tenían correlaciones más fuertes con el veneno de serpiente que las dietas o tipos de presas.[55] Aunque las serpientes venenosas usan sus venenos en defensa (de ahí el problema de la mordedura de serpiente en humanos), no se sabe bien en qué medida la selección natural para la defensa ha impulsado la evolución del veneno. Los venenos de la serpiente coral de Texas, Micrurus tener, y otras especies de Micrurus se ha encontrado que contienen toxinas con actividad específica para inducir dolor, sugiriendo una función defensiva.[56] Sin embargo, una encuesta de cuestionarios a pacientes de mordeduras de serpiente por una amplia variedad de especies venenosas mostró que el dolor después de la mayoría de las mordeduras de serpiente es de inicio lento, argumentando en contra de una selección generalizada para la defensa.[3] El escupir veneno mostrado por algunas especies de cobra escupidora es únicamente una adaptación defensiva. Un estudio de 2021 mostró que los venenos de las tres líneas de cobras escupidoras evolucionaron convergentemente hacia niveles más altos de activación de neuronas sensoriales (es decir, causan más dolor) que los venenos de las cobras no escupidoras, a través de la acción sinérgica de citotoxinas y toxinas fosfolipasa A2, indicando una selección para una función defensiva.[57]
En contraste con la composición del veneno y la toxicidad para linajes específicos, el rendimiento de veneno, o la cantidad de veneno producida por una serpiente individual, no se ha encontrado que varíe con la masa corporal de los animales presa, y en cambio varía con la masa corporal de las serpientes que lo producen. El rendimiento aumenta con la masa corporal de la serpiente de manera consistente con la hipótesis de que las serpientes invierten una proporción constante de la producción metabólica en producir veneno, que es metabólicamente costoso.[47]
Adaptaciones funcionales
.JPG)
Las serpientes usan su veneno para matar o someter a la presa, así como para otras funciones relacionadas con la dieta, como la digestión. La teoría científica actual sugiere que el veneno de serpiente no se usa para la defensa ni para la competencia entre miembros de la misma especie, a diferencia de otros taxones. Por lo tanto, la evolución adaptativa en el veneno de serpiente ha resultado en varias adaptaciones con respecto a estas funciones relacionadas con la dieta que aumentan la aptitud de las serpientes que las portan.[58][43][17] Esto también se refleja en la variación en la composición del veneno dentro de una especie; se sabe que el veneno varía geográficamente, y por edad y tamaño, probablemente reflejando la variación en la presa consumida por diferentes grupos dentro de una especie.[13] La variación geográfica también está presente a nivel de especie; las serpientes de isla tienden a tener venenos menos complejos, mientras que aquellas que viven en hábitats altamente productivos tienen venenos más complejos, sugiriendo un patrón biogeográfico.[59]
Toxicidad del veneno específica para la presa

Se ha encontrado veneno que es tóxico solo para un cierto taxón, o fuertemente tóxico solo para un cierto taxón, en varias serpientes, sugiriendo que estos venenos han evolucionado a través de la selección natural para someter a especies de presas preferidas. Ejemplos de este fenómeno se han encontrado en la serpiente de manglar Boiga dendrophila, que tiene un veneno específicamente tóxico para las aves,[48] así como en los géneros Echis y Sistrurus, y en las serpientes marinas. El veneno de Spilotes sulphureus tiene dos componentes, uno de los cuales es tóxico para los lagartos pero no tóxico en mamíferos, mientras que el otro es tóxico en mamíferos y no tóxico en lagartos.[60] Sin embargo, aunque varias serpientes poseen veneno que es altamente tóxico para sus especies de presas preferidas, la correlación inversa no es necesariamente cierta: los venenos de varias serpientes son tóxicos para taxones que no consumen en grandes proporciones. La mayoría de los venenos de serpiente, por ejemplo, son altamente tóxicos para los lagartos, aunque la proporción de presas de lagartos varía entre las especies de serpientes. Esto ha llevado a los investigadores a sugerir que la toxicidad para un cierto taxón es casi independiente de la toxicidad para otro taxón distantemente relacionado.[41]

Las dietas naturales de las serpientes en el género víbora extendido Echis son altamente variadas, e incluyen artrópodos, como escorpiones, así como vertebrados. Varias especies de Echis consumen diferentes cantidades de artrópodos en su dieta.[61] Un estudio de 2009 inyectó escorpiones con el veneno de varias especies de Echis, y encontró una alta correlación entre la proporción de artrópodos que las serpientes consumían en su hábitat natural, y la toxicidad de su veneno para los escorpiones.[44] Los investigadores también encontraron evidencia de que la evolución de un veneno más tóxico para los artrópodos estaba relacionada con un aumento en la proporción de artrópodos en la dieta de las serpientes, y que la dieta y el veneno podrían haber evolucionado por un proceso de coevolución. Una filogenia del género construida usando ADN mitocondrial mostró que un caso de cambio en la composición del veneno en la especie ancestral de todas las serpientes Echis estaba correlacionado con un cambio a una dieta basada en artrópodos, mientras que otro cambio en un linaje más reciente estaba correlacionado con un cambio a una dieta de vertebrados.[62] A pesar de la mayor toxicidad del veneno de las especies que consumen artrópodos, no se encontró que incapacitara o matara a la presa más rápido que el de las especies con menos artrópodos en su dieta. Por lo tanto, se piensa que el veneno evolucionó para minimizar el volumen requerido, ya que la producción de veneno conlleva un costo metabólico significativo, proporcionando así un beneficio de aptitud.[62] Este patrón también se encuentra en otros linajes.[63] Resultados similares fueron obtenidos por un estudio de 2012 que encontró que el veneno de las especies de Echis que consumen artrópodos era más tóxico para langostas que el de las especies que consumen vertebrados.[64]
Un estudio de 2009 sobre el veneno de cuatro especies de víboras de foseta Sistrurus encontró una variación significativa en la toxicidad para ratones.[41] Esta variación estaba relacionada con la proporción de mamíferos pequeños en la dieta de esas especies.[41] La idea de que el veneno de Sistrurus había evolucionado para adaptarse a una dieta basada en mamíferos fue respaldada por el análisis filogenético. Los investigadores sugirieron que la base para la diferencia en toxicidad era la diferencia en la fisiología muscular en los diversos animales presa.[41] Dos linajes de serpientes elápidos, las serpientes marinas comunes y las Laticauda, han colonizado independientemente ambientes marinos, y han cambiado a una dieta muy simple basada en teleósteos, o peces de aleta radiada.[17] Un estudio de 2005 encontró que ambos linajes tienen un conjunto de proteínas de veneno mucho más simple que sus parientes terrestres en el continente australiano, que tienen una dieta más variada y compleja.[17] Estos hallazgos fueron confirmados por un estudio de 2012, que comparó los venenos de Toxicocalamus longissimus, una especie terrestre, y Hydrophis cyanocinctus, una especie marina, ambas dentro de la subfamilia Hydrophiinae (que también está dentro de la familia Elapidae). A pesar de estar estrechamente relacionadas entre sí, la especie marina tenía un conjunto de proteínas de veneno significativamente más simple.[16] Los venenos de las serpientes marinas son, no obstante, entre los venenos más tóxicos conocidos. Se ha argumentado que, dado que las serpientes marinas típicamente no pueden prevenir la fuga de la presa mordida, sus venenos han evolucionado para actuar muy rápidamente.[65]
Pre-digestión de la presa

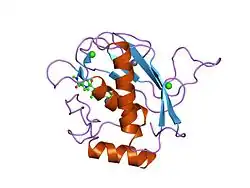
Las diversas subespecies del género de serpientes de cascabel Crotalus producen venenos que cumplen dos funciones conflictivas. El veneno inmoviliza a la presa después de una mordedura, y también ayuda a la digestión al descomponer los tejidos antes de que la serpiente coma a su presa.[18] Como con otros miembros de la familia Viperidae, los venenos de Crotalus interrumpen los procesos homeostáticos de los animales presa. Sin embargo, hay una amplia variedad de composiciones de veneno entre las especies de Crotalus. Un estudio de 2010 encontró una diferencia de 100 veces en la cantidad de actividad de metaloproteinasas entre las diversas serpientes, con Crotalus cerberus teniendo la actividad más alta y Crotalus oreganus concolor teniendo la más baja. También hubo una variación de 15 veces en la cantidad de actividad de proteasa, con C. o. concolor y C. cerberus teniendo las actividades más alta y más baja, respectivamente.[18]
La actividad de las metaloproteinasas causa hemorragia y necrosis después de una mordedura de serpiente, un proceso que ayuda a la digestión. La actividad de las proteasas, por otro lado, interrumpe la función de las plaquetas y los músculos y daña las membranas celulares, y por lo tanto contribuye a una muerte rápida para el animal presa.[18] El estudio encontró que los venenos de Crotalus caían en dos categorías; aquellos que favorecían las metaloproteinasas (Tipo I) y aquellos que favorecían las proteasas (Tipo II). El estudio afirmó que estas funciones eran esencialmente mutuamente excluyentes; los venenos habían sido seleccionados basados en su toxicidad o su potencial de ablandamiento. Los investigadores también plantearon la hipótesis de que la razón para esta dicotomía era que un veneno alto en neurotoxicidad, como un veneno tipo II, mata a un animal rápidamente, impidiendo que la metaloproteinasas de acción relativamente más lenta digiera el tejido.[18]
Rastreo de la presa mordida

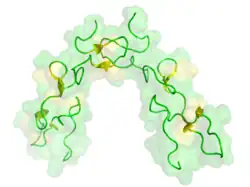
Otro ejemplo de una función adaptativa distinta a la inmovilización de la presa es el papel del veneno de víboras en permitir a la serpiente rastrear a un animal presa que ha mordido, un proceso conocido como "relocalización de la presa." Esta importante adaptación permitió a las serpientes de cascabel evolucionar el mecanismo de mordida y liberación, que proporcionó un enorme beneficio a las serpientes al minimizar el contacto con animales presa potencialmente peligrosos.[19] Sin embargo, esta adaptación luego requiere que la serpiente rastree al animal mordido para comerlo, en un entorno lleno de otros animales de la misma especie. Un estudio de 2013 encontró que las serpientes de cascabel de diamante occidental (Crotalus atrox) respondían más activamente a los cadáveres de ratones que habían sido inyectados con veneno crudo de serpiente de cascabel. Cuando los diversos componentes del veneno fueron separados, las serpientes respondieron a los ratones inyectados con dos tipos de desintegrinas. El estudio concluyó que estas proteínas desintegrinas eran responsables de permitir a las serpientes rastrear a su presa, al cambiar el olor del animal mordido.[19]
Atrofia basada en la dieta

Se cree que el veneno en varias líneas de serpientes se ha atrofiado en respuesta a cambios en la dieta.[7] Un estudio de 2005 en la serpiente marina marmoleada, Aipysurus eydouxii encontró que el gen para una proteína de tres dedos encontrada en el veneno había sufrido una deleción de dos nucleótidos base que hicieron el veneno 50–100 veces menos tóxico de lo que había sido anteriormente. Este cambio estaba correlacionado con un cambio en la dieta de peces a una dieta que consistía casi enteramente en huevos de peces, sugiriendo que la adaptación a una dieta de huevos había eliminado la presión de selección necesaria para mantener un veneno altamente tóxico, permitiendo que los genes de veneno acumularan mutaciones deletéreas.[17][66] Una degradación de veneno similar tras un cambio a una dieta basada en huevos se ha encontrado en el comedor de huevos común Dasypeltis scabra, cuya dieta consiste enteramente en huevos de aves, lo que significa que la serpiente no tenía uso para su veneno. Esto ha llevado a los biólogos a sugerir que si el veneno no es usado por una especie, se pierde rápidamente.[67]
Referencias
- ↑ Hargreaves et al. (a), 2014.
- ↑ Casewell et al., 2013, pp. 218–220.
- ↑ a b Ward-Smith et al., 2020.
- ↑ Fry et al., 2012a, pp. 441–442.
- ↑ a b Wuster et al., 2008.
- ↑ Lomonte et al. (a), 2014, p. 326.
- ↑ a b c d e f Fry et al., 2012a, pp. 434–436.
- ↑ a b Fry et al., 2012a, pp. 424–436.
- ↑ a b Casewell et al., 2013, pp. 224–227.
- ↑ a b c d e f Reyes-Velasco et al., 2015.
- ↑ a b c Hargreaves et al. (b), 2014, pp. 153–155.
- ↑ a b Xie et al., 2022.
- ↑ a b c d e Casewell et al., 2020, pp. 570–581.
- ↑ Casewell et al., 2013, pp. 222–223.
- ↑ Barlow et al., 2009, pp. 2447–2448.
- ↑ a b Calvete et al., 2012, pp. 4094–4098.
- ↑ a b c d e f Li et al., 2005.
- ↑ a b c d e Mackessy, 2010.
- ↑ a b c Saviola et al., 2013.
- ↑ a b Fry et al., 2012a, p. 443.
- ↑ a b Fry et al., 2012a.
- ↑ a b Lomonte et al. (a), 2014, pp. 326–327.
- ↑ Mackessy, 2010, p. 1464.
- ↑ Casewell et al., 2013, pp. 225–227.
- ↑ Palci et al., 2021.
- ↑ Fry et al., 2012a, p. 435.
- ↑ a b Almeida et al., 2021.
- ↑ a b Sunagar y Abraham, 2021.
- ↑ Fry y Wuster, 2004, p. 870.
- ↑ Mount y Brown, 2022, pp. 973–985.
- ↑ Hargreaves et al. (b), 2014.
- ↑ Rao et al., 2022.
- ↑ Casewell et al., 2013, p. 223.
- ↑ a b c Lynch, 2007.
- ↑ a b Dowell et al., 2016.
- ↑ a b Casewell et al., 2013, p. 223–224.
- ↑ Casewell et al., 2011.
- ↑ Fry y Wuster, 2004, p. 871.
- ↑ Fry et al., 2012b.
- ↑ a b c Mikheyev y Barua, 2019.
- ↑ a b c d e f g Gibbs y Mackessy, 2009.
- ↑ Lomonte et al. (a), 2014, p. 334.
- ↑ a b c d Barlow et al., 2009, p. 2443.
- ↑ a b Barlow et al., 2009, p. 2447.
- ↑ Casewell et al., 2013, p. 220.
- ↑ Gibbs y Rossiter, 2008.
- ↑ a b c Healy y Carbone, Jackson, pp. 527–537.
- ↑ a b Casewell et al., 2013, pp. 220–221.
- ↑ Lomonte et al. (b), 2014, pp. 112–114.
- ↑ Heatwole y Poran, 1995.
- ↑ Biardi, Chien y Coss, 2005.
- ↑ Biardi y Coss, 2011.
- ↑ Pomento et al., 2016.
- ↑ Sanz et al., 2006, pp. 2098–2099.
- ↑ Zancolli et al., 2019.
- ↑ Bohlen, 2011.
- ↑ Kazandjian et al., 2021.
- ↑ Casewell et al., 2013, pp. 219–220.
- ↑ Siqueira‐Silva et al., 2021, pp. 1978–1989.
- ↑ Modahl et al., 2018.
- ↑ Barlow et al., 2009, pp. 2444, 2447.
- ↑ a b Barlow et al., 2009, pp. 2446–2448.
- ↑ Casewell et al., 2013, pp. 223–225.
- ↑ Richards et al., 2012.
- ↑ Calvete et al., 2012, pp. 4092–4093.
- ↑ Calvete et al., 2012, pp. 4097–4098.
- ↑ Fry et al., 2008.
Fuentes citadas
- Almeida, Diego Dantas; Viala, Vincent Louis; Nachtigall, Pedro Gabriel; Broe, Michael; Gibbs, H. Lisle; Serrano, Solange Maria de Toledo; Moura-da-Silva, Ana Maria; Ho, Paulo Lee; Nishiyama-Jr, Milton Yutaka; Junqueira-de-Azevedo, Inácio L. M. (10 de mayo de 2021). «Tracking the recruitment and evolution of snake toxins using the evolutionary context provided by the Bothrops jararaca genome» [Seguimiento del reclutamiento y evolución de toxinas de serpiente utilizando el contexto evolutivo proporcionado por el genoma de Bothrops jararaca]. Proceedings of the National Academy of Sciences 118 (20). Bibcode:2021PNAS..11815159A. ISSN 0027-8424. PMC 8157943. PMID 33972420. doi:10.1073/pnas.2015159118. Consultado el 9 de julio de 2025.
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wuster, E.W. (2009). «Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution» [Coevolución de la dieta y la actividad del veneno específica de la presa apoya el papel de la selección en la evolución del veneno de serpiente]. Proceedings of the Royal Society B: Biological Sciences 276 (1666): 2443-2449. PMC 2690460. PMID 19364745. doi:10.1098/rspb.2009.0048. Consultado el 9 de julio de 2025.
- Biardi, James E.; Chien, David C.; Coss, Richard G. (2005). «California ground squirrel (Spermophilus beecheyi) defenses against rattlesnake venom digestive and hemostatic toxins» [Defensas de la ardilla terrestre de California (Spermophilus beecheyi) contra las toxinas digestivas y hemostáticas del veneno de serpiente de cascabel]. Journal of Chemical Ecology 31 (11): 2501-2518. Bibcode:2005JCEco..31.2501B. PMID 16273425. S2CID 23238627. doi:10.1007/s10886-005-7610-1.
- Biardi, JE; Coss, RG (2011). «Rock squirrel (Spermophilus variegatus) blood sera affects proteolytic and hemolytic activities of rattlesnake venoms» [El suero de sangre de la ardilla roquera (Spermophilus variegatus) afecta las actividades proteolíticas y hemolíticas de los venenos de serpientes de cascabel]. Toxicon 57 (2): 323-31. Bibcode:2011Txcn...57..323B. PMID 21184770. doi:10.1016/j.toxicon.2010.12.011.
- Bohlen, Christopher J. (17 de noviembre de 2011). «A heteromeric Texas coral snake toxin targets acid-sensing ion channels to produce pain» [Una toxina heteromérica de la serpiente coral de Texas se dirige a canales iónicos sensibles al ácido para producir dolor]. Nature 479 (7373): 414. Bibcode:2011Natur.479..410B. PMC 3226747. PMID 22094702. doi:10.1038/nature10607. Consultado el 9 de julio de 2025.
- Calvete, J.J.; Ghezellou, P.; Paiva, O.; Matainaho, T.; Ghassempour, A.; Goudarzi, H.; Kraus, F.; Sanz, L. et al. (2012). «Snake venomics of two poorly known Hydrophiinae: Comparative proteomics of the venoms of terrestrial Toxicocalamus longissimus and marine Hydrophis cyanocinctus» [Venómica de serpientes de dos Hydrophiinae poco conocidas: Proteómica comparativa de los venenos de Toxicocalamus longissimus terrestre y Hydrophis cyanocinctus marino]. Journal of Proteomics 75 (13): 4091-4101. PMID 22643073. doi:10.1016/j.jprot.2012.05.026.
- Casewell, N. R.; Wagstaff, S. C.; Harrison, R. A.; Renjifo, C.; Wuster, W. (4 de abril de 2011). «Domain Loss Facilitates Accelerated Evolution and Neofunctionalization of Duplicate Snake Venom Metalloproteinase Toxin Genes» [La pérdida de dominios facilita la evolución acelerada y la neofuncionalización de genes de toxinas metaloproteinasas de veneno de serpiente duplicados]. Molecular Biology and Evolution 28 (9): 2637-2649. ISSN 0737-4038. PMID 21478373. doi:10.1093/molbev/msr091.
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. (2013). «Complex cocktails: the evolutionary novelty of venoms» [Cócteles complejos: la novedad evolutiva de los venenos]. Trends in Ecology & Evolution 28 (4): 219-229. Bibcode:2013TEcoE..28..219C. PMID 23219381. doi:10.1016/j.tree.2012.10.020.
- Casewell, Nicholas R.; Jackson, Timothy N.W.; Laustsen, Andreas H.; Sunagar, Kartik (2020). «Causes and Consequences of Snake Venom Variation» [Causas y consecuencias de la variación del veneno de serpiente]. Trends in Pharmacological Sciences (Elsevier BV) 41 (8): 570-581. ISSN 0165-6147. PMC 7116101. PMID 32564899. doi:10.1016/j.tips.2020.05.006. Consultado el 9 de julio de 2025.
- Dowell, Noah L.; Giorgianni, Matt W.; Kassner, Victoria A.; Selegue, Jane E.; Sanchez, Elda E.; Carroll, Sean B. (2016). «The Deep Origin and Recent Loss of Venom Toxin Genes in Rattlesnakes» [El origen profundo y la pérdida reciente de genes de toxinas de veneno en serpientes de cascabel]. Current Biology 26 (18): 2434-2445. Bibcode:2016CBio...26.2434D. PMC 5207034. PMID 27641771. doi:10.1016/j.cub.2016.07.038. Consultado el 9 de julio de 2025.
- Gibbs, H. Lisle; Rossiter, Wayne (6 de febrero de 2008). «Rapid Evolution by Positive Selection and Gene Gain and Loss: PLA2 Venom Genes in Closely Related Sistrurus Rattlesnakes with Divergent Diets» [Evolución rápida por selección positiva y ganancia y pérdida de genes: Genes de veneno PLA2 en serpientes de cascabel Sistrurus estrechamente relacionadas con dietas divergentes]. Journal of Molecular Evolution 66 (2): 151-166. Bibcode:2008JMolE..66..151G. ISSN 0022-2844. PMID 18253686. S2CID 3733114. doi:10.1007/s00239-008-9067-7.
- Fry, B.G.; Casewell, N.R.; Wuster, W.; Vidal, N.; Young, B.; Jackson, T. N. W. (2012). «The structural and functional diversification of the Toxicofera reptile venom system» [La diversificación estructural y funcional del sistema de veneno de reptiles Toxicofera]. Toxicon 60 (4): 434-448. Bibcode:2012Txcn...60..434F. PMID 22446061. doi:10.1016/j.toxicon.2012.02.013.
- Fry, Bryan G.; Wuster, Wolfgang (2004). «Assembling an Arsenal: Origin and Evolution of the Snake Venom Proteome Inferred from Phylogenetic Analysis of Toxin Sequences» [Ensamblando un arsenal: Origen y evolución del proteoma de veneno de serpiente inferido del análisis filogenético de secuencias de toxinas]. Molecular Biology and Evolution 5 (21): 870-883. PMID 15014162. doi:10.1093/molbev/msh091.
- Fry, Bryan G.; Scheib, Holger; Junqueira de Azevedo, Inacio de L.M.; Silva, Debora Andrade; Casewell, Nicholas R. (2012). «Novel transcripts in the maxillary venom glands of advanced snakes» [Nuevos transcritos en las glándulas de veneno maxilar de serpientes avanzadas]. Toxicon 59 (7–8): 696-708. Bibcode:2012Txcn...59..696F. ISSN 0041-0101. PMID 22465490. doi:10.1016/j.toxicon.2012.03.005.
- Fry, Bryan G.; Scheib, Holger; Van Der Weerd, Louise; Young, Bruce; McNaughtan, Judith; Ramjan, S. F. Ryan; Vidal, Nicolas; Poelmann, Robert E. et al. (2008). «Evolution of an Arsenal» [Evolución de un arsenal]. Molecular & Cellular Proteomics 7 (2): 215-246. PMID 17855442. doi:10.1074/mcp.M700094-MCP200.
- Gibbs, H.L.; Mackessy, S.P. (2009). «Functional basis of a molecular adaptation: Prey-specific toxic effects of venom from Sistrurus rattlesnakes» [Base funcional de una adaptación molecular: Efectos tóxicos específicos de la presa del veneno de serpientes de cascabel Sistrurus]. Toxicon 53 (6): 672-679. Bibcode:2009Txcn...53..672G. PMID 19673082. doi:10.1016/j.toxicon.2009.01.034.
- Hargreaves, Adam D.; Swain, Martin T.; Hegarty, Matthew J.; Logan, Darren W.; Mulley, John F. (1 de agosto de 2014). «Restriction and Recruitment—Gene Duplication and the Origin and Evolution of Snake Venom Toxins» [Restricción y reclutamiento—Duplicación génica y el origen y evolución de las toxinas de veneno de serpiente]. Genome Biology and Evolution 6 (8): 2088-2095. ISSN 1759-6653. PMC 4231632. PMID 25079342. doi:10.1093/gbe/evu166. Consultado el 9 de julio de 2025.
- Hargreaves, Adam D.; Swain, Martin T.; Logan, Darren W.; Mulley, John F. (2014). «Testing the Toxicofera: Comparative transcriptomics casts doubt on the single, early evolution of the reptile venom system» [Probando la Toxicofera: La transcriptómica comparativa pone en duda la evolución única y temprana del sistema de veneno de reptiles]. Toxicon 92: 140-156. Bibcode:2014Txcn...92..140H. PMID 25449103. doi:10.1016/j.toxicon.2014.10.004. Consultado el 9 de julio de 2025.
- Healy, Kevin; Carbone, Chris; Jackson, Andrew L. (7 de enero de 2019). «Snake venom potency and yield are associated with prey-evolution, predator metabolism and habitat structure» [La potencia y el rendimiento del veneno de serpiente están asociados con la evolución de la presa, el metabolismo del depredador y la estructura del hábitat]. Ecology Letters (Wiley) 22 (3): 527-537. Bibcode:2019EcolL..22..527H. ISSN 1461-023X. PMID 30616302. S2CID 58637804. doi:10.1111/ele.13216. hdl:2262/92842.
- Heatwole, Harold; Poran, Naomie S. (1995). «Resistances of sympatric and allopatric eels to sea snake venoms» [Resistencias de anguilas simpátricas y alopátricas a los venenos de serpientes marinas]. Copeia 1995 (1): 136-147. JSTOR 1446808. doi:10.2307/1446808.
- Kazandjian, T. D.; Petras, D.; Robinson, S. D.; van Thiel, J.; Greene, H. W.; Arbuckle, K.; Barlow, A.; Carter, D. A.; Wouters, R. M.; Whiteley, G.; Wagstaff, S. C.; Arias, A. S.; Albulescu, L.-O.; Plettenberg Laing, A.; Hall, C.; Heap, A.; Penrhyn-Lowe, S.; McCabe, C. V.; Ainsworth, S.; da Silva, R. R.; Dorrestein, P. C.; Richardson, M. K.; Gutiérrez, J. M.; Calvete, J. J.; Harrison, R. A.; Vetter, I.; Undheim, E. A. B.; Wüster, W.; Casewell, N. R. (2021). «Convergent evolution of pain-inducing defensive venom components in spitting cobras» [Evolución convergente de componentes de veneno defensivo que inducen dolor en cobras escupidoras]. Science 371 (6527): 386-390. Bibcode:2021Sci...371..386K. ISSN 0036-8075. PMC 7610493. PMID 33479150. S2CID 231666401. doi:10.1126/science.abb9303. Consultado el 9 de julio de 2025.
- Li, M.; Fry, B.G.; Kini, R.M. (2005). «Eggs-only diet: Its implications for the toxin profile changes and ecology of the marbled sea snake (Aipysurus eydouxii)» [Dieta solo de huevos: Sus implicaciones para los cambios en el perfil de toxinas y la ecología de la serpiente marina marmoleada (Aipysurus eydouxii)]. Journal of Molecular Evolution 60 (1): 81-89. Bibcode:2005JMolE..60...81L. PMID 15696370. S2CID 17572816. doi:10.1007/s00239-004-0138-0. Archivado desde el original el 3 de diciembre de 2011. Consultado el 9 de julio de 2025.
- Lomonte, B.; Fernandez, J.; Sanz, L.; Angulo, Y.; Sasa, M.; Gutierrez, J. M.; Calvete, J. J. (2014). «Venomous snakes of Costa Rica: Biological and medical implications of their venom proteomic profiles analyzed through the strategy of snake venomics» [Serpientes venenosas de Costa Rica: Implicaciones biológicas y médicas de sus perfiles proteómicos de veneno analizados a través de la estrategia de venómica de serpientes]. Journal of Proteomics 105: 323-339. PMID 24576642. doi:10.1016/j.jprot.2014.02.020. hdl:10669/11036.
- Lomonte, B.; Tsai, W. C.; Urena-Diaz, J. M.; Sanz, L.; Mora-Obando, D.; Sanchez, E. E.; Fry, B.G.; Gutierrez, J. M.; Gibbs, H. L.; Sovic, M.G.; Calvete, J.J. (2014). «Venomics of New World pit vipers: Genus-wide comparisons of venom proteomes across Agkistrodon» [Venómica de víboras de foseta del Nuevo Mundo: Comparaciones a nivel de género de los proteomas de veneno en Agkistrodon]. Journal of Proteomics 96: 103-116. PMC 4294458. PMID 24211403. doi:10.1016/j.jprot.2013.10.036. Consultado el 9 de julio de 2025.
- Lynch, V.J. (2007). «Inventing an arsenal: adaptive evolution and neofunctionalization of snake venom phospholipase A(2) genes» [Inventando un arsenal: evolución adaptativa y neofuncionalización de los genes de fosfolipasa A(2) de veneno de serpiente]. BMC Evolutionary Biology 7 (1): 2. PMC 1783844. PMID 17233905. doi:10.1186/1471-2148-7-2. Consultado el 9 de julio de 2025.
- Mackessy, Stephen P. (2010). «Evolutionary trends in venom composition in the Western Rattlesnakes (Crotalus viridis sensu lato): Toxicity vs. tenderizers» [Tendencias evolutivas en la composición del veneno en las serpientes de cascabel occidentales (Crotalus viridis sensu lato): Toxicidad vs. ablandadores]. Toxicon 55 (8): 1463-1474. Bibcode:2010Txcn...55.1463M. PMID 20227433. doi:10.1016/j.toxicon.2010.02.028.
- Palci, Alessandro; LeBlanc, Aaron R. H.; Panagiotopoulou, Olga; Cleuren, Silke G. C.; Mehari Abraha, Hyab; Hutchinson, Mark N.; Evans, Alistair R.; Caldwell, Michael W. et al. (11 de agosto de 2021). «Plicidentine and the repeated origins of snake venom fangs» [Plicidentina y los orígenes repetidos de los colmillos de veneno de serpiente]. Proceedings of the Royal Society B: Biological Sciences (The Royal Society) 288 (1956): 20211391. ISSN 0962-8452. PMC 8354744. PMID 34375553. doi:10.1098/rspb.2021.1391. Consultado el 9 de julio de 2025.
- Mikheyev, Alexander S.; Barua, Agneesh (2019). «Many Options, Few Solutions: Over 60 My Snakes Converged on a Few Optimal Venom Formulations» [Muchas opciones, pocas soluciones: Más de 60 millones de años de serpientes convergieron en unas pocas formulaciones óptimas de veneno]. Molecular Biology and Evolution 36 (9): 1964-1974. PMC 6736290. PMID 31220860. doi:10.1093/molbev/msz125. Consultado el 9 de julio de 2025.
- Modahl, Cassandra M.; Mrinalini, null; Frietze, Seth; Mackessy, Stephen P. (2018). «Adaptive evolution of distinct prey-specific toxin genes in rear-fanged snake venom» [Evolución adaptativa de genes de toxinas específicas para presas distintas en el veneno de serpientes de colmillos traseros]. Proceedings of the Royal Society B: Biological Sciences 285 (1884): 20181003. PMC 6111164. PMID 30068680. doi:10.1098/rspb.2018.1003. Consultado el 8 de julio de 2025.
- Mount, Genevieve G; Brown, Jeremy M (22 de marzo de 2022). «Comparing Likelihood Ratios to Understand Genome-Wide Variation in Phylogenetic Support» [Comparación de ratios de verosimilitud para entender la variación en todo el genoma en el soporte filogenético]. Systematic Biology (Oxford University Press (OUP)) 71 (4): 973-985. ISSN 1063-5157. PMID 35323986. doi:10.1093/sysbio/syac014.
- Pomento, AM; Perry, BW; Denton, RD; Gibbs, HL; Holding, ML (2016). «No safety in the trees: Local and species-level adaptation of an arboreal squirrel to the venom of sympatric rattlesnakes» [Sin seguridad en los árboles: Adaptación local y a nivel de especie de una ardilla arbórea al veneno de serpientes de cascabel simpátricas]. Toxicon 118: 149-55. Bibcode:2016Txcn..118..149P. PMID 27158112. doi:10.1016/j.toxicon.2016.05.003.
- Rao, Wei-qiao; Kalogeropoulos, Konstantinos; Allentoft, Morten E; Gopalakrishnan, Shyam; Zhao, Wei-ning; Workman, Christopher T; Knudsen, Cecilie; Jiménez-Mena, Belén; Seneci, Lorenzo; Mousavi-Derazmahalleh, Mahsa; Jenkins, Timothy P; Rivera-de-Torre, Esperanza; Liu, Si-qi; Laustsen, Andreas H (2022). «The rise of genomics in snake venom research: recent advances and future perspectives» [El auge de la genómica en la investigación de veneno de serpiente: avances recientes y perspectivas futuras]. GigaScience (Oxford University Press (OUP)) 11. ISSN 2047-217X. PMC 8975721. PMID 35365832. doi:10.1093/gigascience/giac024. Consultado el 8 de julio de 2025.
- Reyes-Velasco, Jacobo; Card, Daren C.; Andrew, Audra L.; Shaney, Kyle J.; Adams, Richard H.; Schield, Drew R.; Casewell, Nicholas R.; Mackessy, Stephen P. et al. (1 de enero de 2015). «Expression of Venom Gene Homologs in Diverse Python Tissues Suggests a New Model for the Evolution of Snake Venom» [Expresión de homólogos de genes de veneno en diversos tejidos de pitones sugiere un nuevo modelo para la evolución del veneno de serpiente]. Molecular Biology and Evolution (en inglés) 32 (1): 173-183. ISSN 0737-4038. PMID 25338510. doi:10.1093/molbev/msu294.
- Richards, D. P.; Barlow, A.; Wüster, W. (1 de enero de 2012). «Venom lethality and diet: Differential responses of natural prey and model organisms to the venom of the saw-scaled vipers (Echis)» [Letalidad del veneno y dieta: Respuestas diferenciales de presas naturales y organismos modelo al veneno de las víboras de escama aserrada (Echis)]. Toxicon 59 (1): 110-116. Bibcode:2012Txcn...59..110R. PMID 22079297. doi:10.1016/j.toxicon.2011.10.015.
- Sanz, Libia; Gibbs, H. Lisle; Mackessy, Stephen P.; Calvete, Juan J. (2006). «Venom Proteomes of Closely Related Sistrurus rattlesnakes with Divergent Diets» [Proteomas de veneno de serpientes de cascabel Sistrurus estrechamente relacionadas con dietas divergentes]. Journal of Proteome Research 5 (9): 2098-2112. PMID 16944921. doi:10.1021/pr0602500.
- Saviola, A.J.; Chiszar, D.; Busch, C.; Mackessy, S.P. (2013). «Molecular basis for prey relocation in viperid snakes» [Base molecular para la relocalización de presas en serpientes víperidas]. BMC Biology 11 (1): 20. PMC 3635877. PMID 23452837. doi:10.1186/1741-7007-11-20. Consultado el 8 de julio de 2025.
- Siqueira-Silva, Tuany; Lima, Luiz Antônio Gonzaga; Chaves-Silveira, Jônatas; Amado, Talita Ferreira; Naipauer, Julian; Riul, Pablo; Martinez, Pablo Ariel; Sheard, Catherine (27 de julio de 2021). «Ecological and biogeographic processes drive the proteome evolution of snake venom» [Procesos ecológicos y biogeográficos impulsan la evolución del proteoma del veneno de serpiente]. Global Ecology and Biogeography (Wiley) 30 (10): 1978-1989. Bibcode:2021GloEB..30.1978S. ISSN 1466-822X. S2CID 237649145. doi:10.1111/geb.13359. Consultado el 8 de julio de 2025.
- Sunagar, Kartik; Abraham, Siju V (3 de febrero de 2021). «The Curious Case of the "Neurotoxic Skink": Scientific Literature Points to the Absence of Venom in Scincidae» [El curioso caso del "escinco neurotóxico": La literatura científica señala la ausencia de veneno en Scincidae]. Toxins (MDPI AG) 13 (2): 114. ISSN 2072-6651. PMC 7913497. PMID 33546362. doi:10.3390/toxins13020114. Consultado el 8 de julio de 2025.
- Wuster, Wolfgang; Peppin, Lindsay; Pook, Catherine E.; Walker, Daniel E. (2008). «A nesting of vipers: Phylogeny and historical biogeography of the Viperidae (Squamata: Serpentes)» [Un nido de víboras: Filogenia y biogeografía histórica de las Viperidae (Squamata: Serpentes)]. Molecular Phylogenetics and Evolution 49 (2): 445-459. Bibcode:2008MolPE..49..445W. PMID 18804544. doi:10.1016/j.ympev.2008.08.019.
- Ward-Smith, Harry; Arbuckle, Kevin; Naude, Arno; Wüster, Wolfgang (2020). «Fangs for the memories? A survey of pain in snakebite patients does not support a strong role for defense in the evolution of snake venom composition» [¿Colmillos para los recuerdos? Una encuesta de dolor en pacientes de mordeduras de serpiente no respalda un papel fuerte para la defensa en la evolución de la composición del veneno de serpiente]. Toxins 12 (3): 201. PMC 7150919. PMID 32235759. doi:10.3390/toxins12030201. Consultado el 8 de julio de 2025.
- Xie, Bing; Dashevsky, Daniel; Rokyta, Darin; Ghezellou, Parviz; Fathinia, Behzad; Shi, Qiong; Richardson, Michael K.; Fry, Bryan G. (7 de enero de 2022). «Dynamic genetic differentiation drives the widespread structural and functional convergent evolution of snake venom proteinaceous toxins» [La diferenciación genética dinámica impulsa la evolución convergente estructural y funcional generalizada de las toxinas proteínicas del veneno de serpiente]. BMC Biology (Springer Science and Business Media LLC) 20 (1): 4. ISSN 1741-7007. PMC 8742412. PMID 34996434. doi:10.1186/s12915-021-01208-9. Consultado el 8 de julio de 2025.
- Zancolli, Giulia; Calvete, Juan J.; Cardwell, Michael D.; Greene, Harry W.; Hayes, William K.; Hegarty, Matthew J.; Herrmann, Hans-Werner; Holycross, Andrew T.; Lannutti, Dominic I.; Mulley, John F.; Sanz, Libia (13 de marzo de 2019). «When one phenotype is not enough: divergent evolutionary trajectories govern venom variation in a widespread rattlesnake species» [Cuando un fenotipo no es suficiente: Trayectorias evolutivas divergentes gobiernan la variación del veneno en una especie de serpiente de cascabel ampliamente distribuida]. Proceedings of the Royal Society B: Biological Sciences 286 (1898): 20182735. PMC 6458317. PMID 30862287. doi:10.1098/rspb.2018.2735. Consultado el 8 de julio de 2025.
| Control de autoridades |
|
|---|
 Datos: Q19903038
Datos: Q19903038 Multimedia: Evolution of snake venom / Q19903038
Multimedia: Evolution of snake venom / Q19903038